Dezvoltarea embrionară a insectelor începe de obicei cu zdrobirea superficială a oului, care este asociată cu conținutul bogat al ultimului gălbenuș nutritiv. Bastoderma formată pe suprafața oului este mai groasă pe o parte a suprafeței; această îngroșare dă naștere striei germinale și definește partea ventrală a embrionului.
Pliul inelar format de-a lungul marginii benzii germinale, crescând excesiv și crescând împreună deasupra benzii germinale, dă două straturi de membrane germinale - interior (amnion) și exterior (serosa). Din stratul celular al benzii germinale pe partea îndreptată spre gălbenuș,se separă endodermul primar – germenul comun al endodermului și mezodermului. Ulterior, dâra germinativă este segmentată.
Dezvoltarea postembrionară a insectelor este foarte diversă datorită varietății condițiilor de existență a puilor de insecte și a gradului de adaptare a acestora la aceștia. Se pot distinge două tipuri principale de dezvoltare postembrionară: transformarea incompletă, hemimetabolism și transformarea completă, holometabolism. Holometabolismul se caracterizează prin modificări profunde la juvenili, în urma cărora puieții diferă brusc ca structură de insecta adultă (adult). Prin urmare, trecerea de la starea de puieți, adică de la faza larvară la faza imaginară, necesită o restructurare radicală a organelor; aceasta se realizează cu participarea unei faze intermediare între faza larvară și cea imaginară, timp în care puieții rămân latenți; această fază intermediară este pupală. În timpul stării de pupă, organele interne ale insectei fie suferă mai mult sau mai puțină restructurare, fie suferă distrugere completă (histoliză) și sunt reconstruite; în acest din urmă caz, formarea din nou a organului are loc datorită așa-numitelor discuri imaginare, formate din celule de natură embrionară (histoblaste). Astfel, perioada pupală este o perioadă de histoliză și histogeneză activă (neoplasme tisulare). În exterior, pupa reprezintă faza imaginară cu organele sale caracteristice, care, totuși, au rămas subdezvoltate.
Dezvoltarea la ordinele superioare de insecte este holometabolică la gândaci, lepidoptere, muștele, purici, himenoptere și diptere.
În cazul transformării incomplete - hemimetabolism - nu există o diferență atât de profundă în structura fazelor tinere și a adultului: juvenil acum mai mult, apoi ceva mai puțin seamănă cu faza imaginară ca structură. În sfârșit, în cazurile în care trecerea de la tineret. faza la adult se reduce doar la creșterea și vărsarea (în legătură cu creșterea) vechiului înveliș (exuvia), se vorbește despre amebolism, adică absența transformării; un exemplu de amebolism este dezvoltarea Collembola printre Apterygota. Cu toate acestea, de obicei, cu stadii succesive, care sunt separate între ele prin moarte, adică prin îndepărtarea învelișului epocii anterioare, apar și modificări ale formei organelor: de exemplu, lungimea antenelor și a cercului. crește relativ, numărul de segmente crește; unele organe pot reapărea, cum ar fi anexele genitale externe. Aceasta este dezvoltarea în unele Apterygota, cum ar fi Thysanura și Diplura, și în majoritatea Pterygota hemimetabolice. La unii Pterygota care se dezvoltă cu hemimetabolism (libelule, efee; muște de piatră și părți de artropode), dezvoltarea este complicată din cauza faptului că puii trăiesc în alte condiții decât adulții; astfel, puii de libelule, efee și muștele de piatră se dezvoltă în corpurile de apă și, în legătură cu aceasta, dobândesc organe de respirație a apei - branhii, care sunt un exemplu de organe larvare (provizorii). Condițiile de existență se reflectă și în dezvoltarea altor organe ale larvei, care se adaptează mediului, de exemplu, transformarea picioarelor din față ale cicadei juvenile într-un organ de săpat în legătură cu viața sa subterană.
Dezvoltarea insectelor
Dezvoltarea individuală a insectelor (ontogenia) constă în dezvoltarea embrionară care are loc în faza de ou, iar dezvoltarea postembrionară - după ce larva părăsește oul până la atingerea fazei de adult - imago.
Dezvoltare embrionară. Ouăle de insecte variază ca formă datorită adaptărilor la mediul în care se dezvoltă. De exemplu, la gândaci, ouăle sunt predominant ovale și se dezvoltă mai des într-un substrat închis; în ploșnițe - în formă de butoi, atașat de substrat; la fluturi - în formă de turelă sau în formă de sticlă; în aripioare (ochi aurii) ouă cu tulpină. Ouăle sunt adesea depuse în grupuri. Punctele de ouă pot fi deschise sau închise. Un exemplu de ouat în aer liber sunt ouăle gândacului de cartofi de Colorado, lipite de femelă pe partea inferioară a frunzelor de cartof. Ambreiaje închise includ păstăi de ouă de acridoide formate din particule de sol cimentate de excrețiile glandelor anexe ale femelelor. Gândacii își depun ouăle în ootheca - capsule de ouă care se formează în tractul genital feminin.
Ouăle de insecte sunt acoperite la exterior cu o coajă - corion, care le protejează de uscare (Fig. 337). Pe suprafața cochiliei există un micropil - o mică gaură cu un "dop" complex cu un tub în interior pentru penetrarea spermatozoizilor în timpul fertilizării. Sub corion este o membrană subțire de gălbenuș, iar sub ea este un strat dens de citoplasmă. Partea centrală a citoplasmei este umplută cu gălbenuș. Citoplasma conține nucleul și corpurile polare.
Zdrobirea este superficială. Inițial, nucleul se divide de mai multe ori, nucleii fiice cu secțiuni ale citoplasmei migrează la periferia oului, sunt acoperiți cu o membrană și se formează un strat de suprafață de celule - blastodermul, iar gălbenușul rămâne în centrul oului. ou. Pe suprafața ventrală a blastodermului, celulele sunt mai înalte și formează o îngroșare - banda germinativă. Acest stadiu al dezvoltării embrionare a insectelor corespunde blastulei.
Diviziunea celulară a benzii germinale duce la dezvoltarea embrionului. Fâșia germinativă se scufundă treptat, formând șanțul ventral.
Orez. 337. Structura unui ou de insectă (din Bei-Bienko): 1 - micropil, 2 - corion, 3 - membrana gălbenușă, 4 - nucleu, 5 - corpi polari, 6 - gălbenuș

Pliurile blastodermei de deasupra brazdei se închid și se formează membrane germinale: seroasă și amnios (Fig. 338). Aici se manifestă convergența cu vertebratele superioare, care au și cochilii asemănătoare. Datorită cavității amniotice formate, embrionul se află într-o stare suspendată în interiorul oului, ceea ce îl protejează în mod fiabil de deteriorarea mecanică.În plus, lichidul care umple cavitatea amniotică facilitează procesele metabolice ale embrionului.
Banda germinativă se diferențiază în continuare în două straturi: inferior - ectoderm și superior - endomezoderm. Entomezodermul la diferite specii de insecte se poate forma în diferite moduri: prin invaginare sau prin imigrare celulară.
În următoarea etapă de dezvoltare, stratul ectodermic al benzii începe să se îndoaie în sus pe laterale, apoi se închide pe spate, formând un perete închis al embrionului. Când pereții corpului se închid pe spate, o parte din gălbenușul și celulele gălbenușului intră în corpul embrionului. Concomitent cu formarea pereților corpului embrionului, două grupuri de celule sunt separate în entomezoderm la capetele anterioare și posterioare ale corpului. Acestea sunt cele două rudimente ale intestinului mediu. Ulterior, secțiunile anterioare și posterioare ale intestinului mediu încep să se formeze din aceste două primordii, care apoi fuzionează. În același timp, la capetele anterioare și posterioare ale corpului embrionului se formează invaginări profunde ale ectodermului, din care se formează secțiunile anterioare și posterioare ale intestinului. Apoi toate cele trei secțiuni sunt conectate, formând un tub intestinal traversant.
Dâra mezodermică se descompune în rudimente metamerice pereche ale sacilor celomici. Dar mai târziu se dezintegrează, iar din mezoderm se formează mușchii embrionului, foaia somatică a epiteliului celomic, inima, corpul gras și gonadele. Stratul visceral al epiteliului celomic nu se formează la insecte, iar cavitatea corpului devine amestecată - mixocoelul. Rudimentele celomice se contopesc cu cavitatea primară a corpului.
Mai târziu, din ectoderm se formează sistemul nervos și sistemul traheal. Vasele malpighiene se formează din pereții intestinului posterior.
În procesul de dezvoltare, embrionul de insectă suferă o segmentare, care se manifestă mai întâi în partea anterioară, apoi în partea posterioară a corpului. În secțiunea capului, un acron cu ochi, labial și

lobi antene, un segment intercalar și trei segmente maxilare. Apoi se formează trei segmente toracice și zece abdominale și un lob anal.
La multe insecte, embrionul trece prin trei etape, caracterizate printr-o compoziție diferită a primordiilor membrelor: protopod, polipod și oligopod (Fig. 339).
Dezvoltarea embrionară a insectelor se caracterizează prin fenomenul de blastokineză. Aceasta este o schimbare a poziției corpului embrionului în ou, în care rezervele de gălbenuș sunt cele mai utilizate.
Două tipuri de blastokineză pentru insecte sunt descrise de A. G. Sharov. La insectele cu transformare incompletă, la început embrionul este situat cu spatele în sus și capul spre capătul anterior al oului, iar apoi, când se formează cavitatea amniotică, embrionul se întoarce cu partea ventrală în sus, iar capul, respectiv, se află în spatele oului.
Blastokineza apare diferit la majoritatea insectelor cu transformare completă și la ortoptere, în care embrionul se scufundă în gălbenuș fără a schimba poziția corpului în ou.
În dezvoltarea embrionară a insectelor se manifestă adaptări la viața de pe uscat: membrane protectoare (corion, serosa, amnion), o aport de nutrienți (mult gălbenuș), o cavitate amniotică plină cu lichid.
Înainte de eclozare, larva de insectă formată înghite lichidul din cavitatea amniotică, datorită căruia turgul corpului crește. Larva sparge corionul cu capul, care are adesea dinți de ou sau un vârf.
Dezvoltare postembrionară. În perioada de dezvoltare postembrionară a insectelor, după ecloziunea din ou, creșterea unui animal tânăr are loc prin mușcări succesive și trecerea unor faze de dezvoltare calitativ diferite. În timpul ontogenezei sau dezvoltării individuale, insectele năpădesc de la 3-4 la 30 de ori. În medie, numărul de link-uri este de 5-6. Intervalul dintre moarte se numește stadiu, iar starea de dezvoltare se numește vârstă. Modificările morfologice din timpul dezvoltării de la larvă la adult se numesc metamorfoză. La toate insectele, cu excepția formelor inferioare fără aripi, după atingerea stării adulte, creșterea și năpârlirea adulților încetează. Prin urmare, de exemplu, variațiile în dimensiunea gândacilor din aceeași specie nu pot fi atribuite diferitelor grupe de vârstă, ci ar trebui considerate doar o manifestare a variabilității individuale.
Există trei tipuri principale de dezvoltare postembrionară a insectelor: 1) dezvoltare directă fără metamorfoză - ametabolism, sau protometabolism; 2) dezvoltare cu transformare incompletă, sau cu metamorfoză treptată, - hemimetabolism; 3) dezvoltare cu transformare completă, i.e. cu o metamorfoză pronunțată – holometabolism.
Ametabolism, sau dezvoltare directă, se observă numai la insectele primare fără aripi din ordinul cozii cu peri (Thysanura), care includ peștele de argint comun (Lepisma). Același tip de dezvoltare se observă și la Entognatha: coada arcului (Collembola) și cu două cozi (Diplura).
Cu ametabolism, o larvă, asemănătoare unui adult, iese din ou. Diferentele privesc doar marimea, proportiile corpului si gradul de dezvoltare al gonadelor. Spre deosebire de insectele înaripate, ele continuă să naparească chiar și în starea imaginară.
Hemimetabolism- transformare incompletă, sau dezvoltare cu metamorfoză treptată. Este caracteristic multor insecte înaripate, de exemplu, gândaci, lăcuste, lăcuste, ploșnițe, cicade etc.
Cu hemimetabolism, din ou iese o larvă, asemănătoare adulților, dar cu aripi rudimentare și gonade subdezvoltate. Astfel de larve asemănătoare adulților cu rudimente de aripi se numesc nimfe. Acest nume este împrumutat din mitologia greacă veche și se referă la ființe divine înaripate sub formă de fete. Nimfele insectelor năpardesc de mai multe ori, iar cu fiecare năpârlire, rudimentele aripilor lor cresc. O nimfă mai în vârstă se mută și din ea iese o imagine înaripată. Figura 340 prezintă fazele dezvoltării lăcustelor (ouă, nimfe de stadii 1-5 și adulți) ca exemplu de transformare incompletă. Această transformare incompletă tipică se numește hemimetamorfoză.
Printre insectele cu metamorfoză incompletă, există cazuri de dezvoltare când nimfele diferă semnificativ de adulți în prezența unor adaptări speciale larvare - organe provizorii. O astfel de dezvoltare se observă la libelule, efei, muștele de piatră. Nimfele acestor insecte trăiesc în apă și, prin urmare, sunt numite

Orez. 340. Dezvoltare cu metamorfoză incompletă la lăcusta Locusta migratoria (după Kholodkovsky): 1 - protorax, 2 - mezotorax cu muguri de aripi, 3 - metatorax cu muguri de aripi
naiade (nimfe de apă). Au organe provizorii precum branhiile traheale, care dispar la adulții de pe uscat. Și larvele de libelule au încă o „mască” - o buză inferioară modificată care servește la prinderea pradă.
Holometabolism- transformare completă. Fazele de dezvoltare în holometabol sunt: ou - larvă - pupă - imago (Fig. 341). Această dezvoltare este tipică pentru gândaci, fluturi, diptere, himenoptere, caddisflies și lacewings.

Orez. 341. Dezvoltare cu transformare completă în vârful de vierme de mătase Bombyx (după Leines): A - mascul, B - femela, C - omida, D - cocon, E - pupă de cocon
Larvele de insecte cu metamorfoză completă nu sunt asemănătoare cu adulții și adesea diferă din punct de vedere ecologic. Deci, de exemplu, larvele gândacilor de mai trăiesc în sol, iar adulții trăiesc pe copaci. Larvele multor muște se dezvoltă în sol, substrat putrezitor, iar adulții zboară și vizitează florile, hrănindu-se cu nectar. Larvele unor astfel de insecte năpârșesc de mai multe ori și apoi se transformă într-o pupă. În faza pupală are loc histoliza - distrugerea organelor larvare și histogeneza - formarea organizării unei insecte adulte. Din pupă iese o insectă înaripată - o imago.
Astfel, la insecte se observă următoarele tipuri de dezvoltare postembrionară: ametabolism, sau protomorfoză (ou - larvă (asemănătoare cu imago) - imago); hemimetabolie - transformare incompletă (ou - nimfă - imago): hemimetamorfoză - o variantă tipică, hipomorfoză - metamorfoză redusă, hipermorfoză - metamorfoză crescută; holometabol - transformare completă (ou - larvă - pupă - adult): holometamorfoză - o variantă tipică, hipermetamorfoză - cu mai multe tipuri de larve.
Tipuri de larve de insecte cu metamorfoză completă. Larvele insectelor holometabolice au o structură mai simplificată în comparație cu adulții. Nu au ochi compuși, rudimente de aripi; aparatul bucal este de tip roadă, antenele și picioarele sunt scurte. În funcție de dezvoltarea membrelor, se disting patru tipuri de larve: protopode, oligopode, polipode și apoduri(Fig. 342). Larvele de protopode sunt caracteristice albinelor, viespilor. Au doar începuturile picioarelor pectorale. Aceste larve sunt inactive, se dezvoltă în faguri cu grijă de la muncitori. Larvele de oligopode sunt mai frecvente decât altele; ele se caracterizează prin dezvoltarea normală a trei perechi de picioare. Oligopodele includ larve de gândaci, aripioare. Larvele polipode, sau omizile, au, pe lângă trei perechi de picioare toracice, mai multe perechi de picioare false pe abdomen. Crura ventrală reprezintă proeminențe ale abdominale
pereții corpului și cârligele de urs și tepii pe talpă. Omizile sunt caracteristice fluturilor și fluturii.Larvele apodice, sau fără picioare, sunt observate în ordinea Diptera, precum și la unii gândaci (larve de mreane, forători), fluturi.

Orez. 342. Larve de insecte cu metamorfoză completă (din Barnes): A - protopod, B, C - oligopod, D - polipod, E, F, G - apod
Conform metodelor de mișcare, larvele de insecte cu metamorfoză completă sunt împărțite în campodeoid cu un corp lung și flexibil, picioare alergătoare și senzație de cerc, erucoid cu un corp cărnos, ușor curbat, cu sau fără membre, sârmă- cu corp rigid, rotund ca diametru, cu cerci de sustinere - urogomphs si asemănător viermilor- fără picioare.
Larvele campodeoide sunt caracteristice multor gândaci prădători - gândaci de pământ, gândaci rove. Se deplasează prin găurile din sol. O larvă tipică erucoidă este larva gândacului de mai, gândacul de bălegar, bronzul. Acestea sunt larve de vizuini. Viermii de sârmă sunt caracteristici gândacilor clic și gândacilor întunecați, ale căror larve fac în mod activ treceri în sol. Numeroase larve asemănătoare viermilor. Se mișcă în sol, în țesuturile plantelor. Acestea includ nu numai larvele de Diptera, ci și câțiva gândaci, fluturi, mușchii care se dezvoltă, de exemplu, în țesuturile plantelor.
tipuri de pupe. Pupele sunt libere, acoperite și ascunse (Fig. 343). În pupele libere, rudimentele aripilor și ale membrelor sunt clar vizibile și separate liber de corp, de exemplu, la gândaci. În pupele acoperite, toate rudimentele aderă strâns la corp, de exemplu, la fluturi. Tegumentele pupelor libere sunt subțiri, moi, în timp ce cele ale pupelor acoperite sunt puternic sclerotizate.

Orez. 343. Tipuri de pupe la insecte (de la Weber): A - gândac liber, B - fluture acoperit, C - muscă ascunsă; 1 - antenă, 2 - rudimente de aripi, 3 - picior, 4 - spiraculi
piele larvară, care formează un fals cocon - puparia. În interiorul pupariumului este o pupă deschisă. Prin urmare, crisalida ascunsă este doar o variantă a celei libere. Pupariile sunt caracteristice multor muște.
Adesea, larva ultimului stadiu țese un cocon înainte de pupație. Deci, de exemplu, o omidă de fluturi de viermi de mătase secretă mătase din glandele de mătase, din care învârte un cocon dens. În interiorul unui astfel de cocon se află o crisalidă acoperită. Și la unele himenoptere - furnici, precum și la lacewings, există o pupă deschisă sau liberă în interiorul coconului. La larvele de aripioare, cum ar fi ochii aurii, firele pentru cocon sunt produse de vasele malpighiene și excretate din anus.
Fiziologia metamorfozei. În procesul de metamorfoză au loc două procese interdependente: histoliza și histogenia. Histoliza este defalcarea țesuturilor organelor larvare, iar histoliza este formarea organelor unei insecte adulte. La insectele cu metamorfoză incompletă, aceste procese au loc treptat în faza de nimfă, iar la insectele cu metamorfoză completă, în faza de pupă.
Histoliza are loc datorită activității fagocitelor și a enzimelor. În acest caz, în primul rând, corpul adipos, mușchii larvarilor și alte câteva organe sunt distruse, care sunt transformate într-un substrat nutritiv consumat de țesuturile în curs de dezvoltare.
Histogenia, sau formarea organelor unei insecte adulte, are loc în principal datorită dezvoltării discurilor imaginare - rudimente din celulele nediferențiate. Discurile imaginare sunt depuse în timpul fazei larvare și chiar în timpul embriogenezei și reprezintă rudimente interne. Discurile imaginare dezvoltă ochii, aripile, aparatul bucal, picioarele, precum și organele interne: mușchi, gonade. Sistemul digestiv, vasele malpighiene și traheele nu sunt distruse, dar sunt puternic diferențiate în procesul de metamorfoză. Cel mai puțin metamorfozat inimă și sistem nervos. Cu toate acestea, în timpul metamorfozei în sistemul nervos, se observă adesea procesul de oligomerizare (fuziune) a ganglionilor.
Procesul de metamorfoză este controlat de glandele endocrine (Fig. 329). celule neurosecretoare creierul secretă hormoni care stimulează activitatea corpuri cardiace, ai caror hormoni prin hemolimfa stimuleaza protoracică glande (protoracice) care secretă hormonul de napârlire - ecdizonă. Ecdysone favorizează procesul de năpârlire: dizolvarea parțială și exfolierea cuticulei vechi, precum și formarea uneia noi.
În procesul de metamorfoză, activitatea corpurile adiacente producând juvenile hormon. La concentrații mari, năpârlirea larvei duce la formarea unei larve
următoarea vârstă. Pe măsură ce larvele cresc, activitatea corpurilor adiacente slăbește și concentrația de hormon juvenil scade, iar glandele protoracice degenerează treptat. Acest lucru duce la faptul că larvele năparesc în faza pupală și apoi adult.
Transplantul artificial al corpurilor adiacente, de exemplu, într-o nimfă de lăcuste din ultima vârstă, contribuie la faptul că aceasta nu se mută în faza adultă, ci într-o larvă mai mare de vârstă suplimentară. În faza adultă, hormonul juvenil controlează dezvoltarea gonadelor, iar hormonul ecdizonă nu se mai produce din cauza reducerii glandelor protoracice.
Originea metamorfozei. Există mai multe ipoteze despre originea metamorfozei la insecte. Multă vreme au existat dispute despre care insecte sunt mai avansate evolutiv - cu transformare completă sau incompletă. Pe de o parte, nimfele de insecte cu metamorfoză incompletă sunt dezvoltate mai progresiv decât larvele de insecte cu metamorfoză completă, pe de altă parte, acestea din urmă au o fază pupală avansată.
În prezent, această contradicție a fost înlăturată de ipoteza originii metamorfozei de către G. S. Gilyarov, A. A. Zakhvatkin și A. G. Sharov. Conform acestei ipoteze, ambele forme de metamorfoză la insecte s-au dezvoltat independent de un tip mai simplu de dezvoltare - protomorfoza, observată la insectele primare fără aripi, de exemplu, la insectele cu coadă cu peri (Thysanura).
În cazul protomorfozei, dezvoltarea este directă, observându-se multe mucegai în faza larvară, iar apoi în stare imaginară. Toate fazele de dezvoltare ale acestor insecte au loc în același mediu.
Se presupune că, în procesul de evoluție, insectele au trecut de la o existență semi-ascunsă în stratul superior al solului la viața la suprafața sa și pe plante. Această tranziție către un nou habitat a fost încununată cu o aromorfoză majoră - dezvoltarea aripilor și a zborului.
Dezvoltarea habitatelor deschise s-a reflectat în evoluția dezvoltării individuale a insectelor. Evoluția ontogenezei insectelor a urmat aparent două direcții principale.
Într-un caz, procesul de embrionizare a dezvoltării era în curs, ducând la ecloziunea insectelor din ouă bogate în gălbenuș în fazele ulterioare de dezvoltare. Acest lucru a dus la imaginarea larvelor cu formarea de nimfe. Așa s-au dezvoltat insectele cu metamorfoză incompletă. Această cale de evoluție a dus la dezvoltarea progresivă a larvelor care conduc un mod de viață similar cu adulții.
În celălalt caz, dimpotrivă, a avut loc un proces de deembrionizare a dezvoltării, adică ouăle sărace în gălbenuș au fost clocite în stadiile anterioare de dezvoltare. Acest lucru a dus la divergența morfoecologică a larvelor de insecte și a adulților. Larvele s-au simplificat și adaptat
să trăiască într-un mediu mai protejat, îndeplinind funcția de nutriție, iar adulții au început să îndeplinească în principal funcția de reproducere și așezare. Pe lângă deembrionizarea dezvoltării larvelor de insecte cu transformare completă, acestea au dezvoltat multe adaptări provizorii la diverse condiții de existență. Astfel, amfigeneza (divergența) s-a produs în evoluția larvelor și a adulților la insectele cu metamorfoză completă. Amfigeneza larvelor și adulților s-a dovedit a fi foarte profundă în ceea ce privește adaptările morfologice, ceea ce a creat contradicții serioase în ontogenie. Ele au fost rezolvate cu succes prin apariția fazei pupale, în timpul căreia are loc o restructurare radicală a organizării larvare la imaginal. Acest lucru a permis insectelor cu transformare completă să stăpânească o gamă mai largă de nișe ecologice și să obțină o înflorire fără precedent printre animalele de pe Pământ.
Creșterea insectelor. Pentru majoritatea insectelor, reproducerea sexuală bisexuală este caracteristică. Multe specii prezintă dimorfism sexual. De exemplu, gândacii de cerb masculi au mandibule modificate în coarne, iar gândacii rinoceri masculi au un corn pe cap și cocoașe pe pronot. Acest lucru se datorează comportamentului de împerechere al acestor specii, însoțit de lupta masculilor pentru o femelă. Relația dintre sexe la diferite specii este extrem de diversă. Dipterele masculi din familia Dolichopodidae aduc un „cadou” femelei - o muscă prinsă și dansează cu oglinzi pentru picioare. Femelele mantis au o dispoziție de prădător și mănâncă masculul în timpul împerecherii.
Majoritatea insectelor depun ouă, dar se observă adesea nașteri vii. În acest caz, ouăle se dezvoltă în tractul genital feminin și ea dă naștere larvelor. De exemplu, sarcofagii (Sarcophagidae) depun larve vii pe carne, a căror dezvoltare se desfășoară foarte repede. Nu e de mirare în antichitate se credea că viermii din carne se generau spontan. Este posibil să nu observați cum o muscă a vizitat carnea întinsă în mod deschis și să descoperiți brusc larve albe care au apărut brusc.
Printre speciile vivipare se numără și musca oi, câțiva gândaci care trăiesc în peșteri.
Pe lângă reproducerea sexuală bisexuală, o serie de insecte prezintă partenogeneză - dezvoltare fără fertilizare. Există multe specii din diferite ordine de insecte, care se caracterizează prin partenogeneză. Partenogeneza poate fi obligatorie - obligatorie, atunci toți indivizii speciei sunt doar femele. Deci, în condiții alpine, în nord și în alte condiții nefavorabile, se găsesc gândaci partenogenetici, ortoptere, urechi și lacewings. Partenogeneza are loc și la speciile bisexuale, când o parte din ouă sunt depuse fertilizate, iar o parte fără fertilizare. De exemplu, trântorii de albine se dezvoltă din ouă nefertilizate.
Partenogeneză similară se găsește și la alte himenoptere (furnici, muștele de ferăstrău), termite, unele gândaci și gândaci. Și la afide, de exemplu, există o schimbare a generațiilor în ciclul de viață: bisexual și partenogenetic. În unele cazuri, partenogeneza poate fi opțională (temporară), manifestându-se numai în condiții nefavorabile. Partenogeneza la insecte contribuie la menținerea populațiilor mari.
O variantă a partenogenezei este pedogeneza - reproducerea fără fertilizare în faza de dezvoltare larvară. Acesta este un mod special de dezvoltare a insectelor, atunci când maturizarea gonadelor este înaintea altor organe. De exemplu, unele specii de muschi biliari se reproduc în faza larvară. Larvele mai bătrâne dau naștere larvelor mai tinere. Pedogeneza a fost remarcată pentru una dintre speciile de gândaci, ale căror larve depun parțial ouă și parțial dau naștere larvelor. Pedogeneza, pe lângă reproducerea bisexuală, crește abundența speciei.
Ciclurile de viață ale insectelor. Spre deosebire de ontogeneză, sau de dezvoltarea individuală a insectelor, ciclul de viață este dezvoltarea unei specii, care, de regulă, include mai multe tipuri de ontogene. Ontogenia se limitează la viața unui individ, de la ovul până la debutul pubertății și apoi moartea naturală. Ciclul de viață este o parte recurentă a dezvoltării continue a unei specii. Astfel, în cel mai tipic caz la insecte, ciclul de viață este format din două ontogene conjugate și morfofiziologic diferite ale masculilor și femelelor, care se reproduc sexual și reproduc propriul lor fel. Și la speciile partenogenetice, ciclul de viață este caracterizat de un singur tip de ontogeneză feminină.
Ciclurile de viață ale insectelor sunt diverse în ceea ce privește tipurile de reproducere, compoziția generațiilor și alternanța lor. Se pot distinge următoarele tipuri de cicluri de viață ale insectelor.
1. Cicluri de viață fără alternanță de generații cu reproducere sexuală bisexuală. Acesta este cel mai frecvent tip de ciclu de viață, caracteristic speciilor dimorfe, format doar din masculi și femele care se reproduc sexual. Acestea sunt ciclurile majorității gândacilor, fluturilor, ploșnițelor.
2. Cicluri de viață fără alternarea generațiilor cu reproducere partenogenetică. Astfel de specii sunt monomorfe, formate doar din femele partenogenetice care depun ouă fără fertilizare.
Speciile partenogenetice sunt deosebit de comune printre afide, psilide și alte homoptere. Speciile partenogenetice de gândaci, gândaci, lăcuste și coccizi sunt comune în condiții de munte înaltă.
3. Cel mai rar tip de ciclu de viață la insecte este un ciclu fără alternanță de generații cu reproducerea sexuală a speciilor hermafrodite.
Este cunoscută o specie americană de muscă, formată doar din indivizi hermafrodiți. În fazele timpurii de dezvoltare, adulții funcționează ca bărbați, iar în etapele ulterioare, ca femele. Prin urmare, toți indivizii depun ouă, ceea ce crește abundența speciei.
4. Cicluri de viață fără alternanță de generație cu reproducere sexuală și partenogeneză facultativă la speciile polimorfe, de exemplu, la insectele sociale. Specia este formată din indivizi sexuali - masculi și femele și fertile - indivizi care lucrează care nu participă la reproducere. Aceste specii includ albine, furnici și termite. Astfel de cicluri de viață sunt complicate de faptul că femelele depun, împreună cu ouăle fecundate, și cele partenogenetice, din care, de exemplu, albinele dezvoltă masculi haploizi - trântori, iar din cele fertilizate - femele și muncitoare. Unii călăreți, trips, coccizi se dezvoltă similar.
La alte specii, partenogeneza facultativă se manifestă într-un mod diferit: nu masculii, ci femelele se dezvoltă din ouă nefertilizate. Dar în acest caz, setul diploid de cromozomi este restaurat la femele prin fuziunea nucleelor haploide. O astfel de dezvoltare este cunoscută la unele insecte stick, lăcuste, muste, coccide.
5. Cicluri de viață cu alternanță de generație sexuală și partenogenetică (heterogonie). La multe afide și filoxere, pe lângă generația sexuală a masculilor și femelelor înaripate, există mai multe generații alternative de femele partenogenetice, înaripate sau fără aripi.
6. Cicluri de viață cu alternanță de generație sexuală și mai multe generații cu pedogeneză. De exemplu, la unele mușchii biliari, după reproducerea sexuală, la care participă masculi și femele, are loc reproducerea partenogenetică a larvelor (pedogeneză). După mai multe generații de larve de reproducție, care mor după nașterea vie a propriului soi, ultima generație de larve se pupează și dă naștere la femele și masculi înaripați.
7. Cicluri de viață cu alternanță de generație sexuală (masculi și femele) cu asexuată. După reproducerea sexuală, femelele depun ouă fertilizate care suferă poliembrionare. Aceasta este reproducerea asexuată în faza embrionară. Oul este supus zdrobirii, iar embrionul în faza de morula începe să se înmulțească prin înmugurire. Dintr-un ou se pot forma câteva zeci de embrioni. Astfel de
Astfel, clasificarea ciclurilor de viață poate fi reprezentată astfel.
eu. Fără alternanță de generații:
- 1) cu reproducere sexuală bisexuală (Gândacul de mai);
- 2) cu reproducere partenogenetică (gândaci alpini, lăcuste);
- 3) cu reproducerea sexuală a indivizilor hermafrodiți (musca americană);
- 4) cu reproducere sexuală și partenogeneză parțială la speciile polimorfe (albine).
II. Cu alternanță de generații:
- 1) heterogonie: alternarea generației sexuale și a mai multor partenogenetice (afidele, filoxera);
- 2) heterogonie: alternarea generaţiei sexuale şi a mai multor generaţii pedogenetice (unii muschi biliari);
- 3) metageneza: alternarea generației sexuale cu poliembrionul (călăreți).
Ciclurile sezoniere ale insectelor. Dacă ciclul de viață este înțeles ca o parte care se repetă ciclic a morfogenezei unei specii de la o fază de dezvoltare la aceeași fază, atunci ciclul de dezvoltare sezonieră este înțeles ca o caracteristică a dezvoltării unei specii în timpul anotimpurilor unui an ( de la iarnă la iarnă).
Deci, de exemplu, ciclul de viață al gândacului de mai durează 4-5 ani (de la ou la adulții maturi), iar ciclul sezonier al acestei specii se caracterizează prin faptul că larvele iernate se pupă primăvara și gândacii tineri se reproduc. Vara, toamna și iarna se găsesc larvele lor de diferite vârste. Numărul de generații care se dezvoltă pe parcursul anului se numește voltness.
Există specii care dau mai multe generații pe an. Acestea sunt tipuri multivoltaice. De exemplu, o musca de casa poate da 2-3 generatii pe sezon si hiberneaza in faza adulta. Majoritatea insectelor sunt monovoltine, producând o generație pe an.
Ciclurile sezoniere ale insectelor din natură sunt caracterizate de perioade calendaristice de apariție a diferitelor faze de dezvoltare. Trăsăturile importante ale ciclurilor sezoniere ale speciilor sunt momentul vieții lor active și diapauza (întârziere temporară a dezvoltării) iarna sau vara. Reglarea ciclurilor de viață ale speciilor în conformitate cu fenomenele sezoniere locale este asigurată de factorii de mediu și de sistemul neuroumoral al organismului.
Dezvoltarea embrionară sau embrionară a albinei lucrătoare, uterului, dronei include toate modificările care au loc sub coaja oului, în urma cărora dintr-un ou unicelular se formează o creatură multicelulară, larva (Fig. 32). ). Oul tocmai depus de uter, de regulă, este lipit de fundul celulei cu un capăt și stă vertical pe el. Lungimea oului este de 1,6-1,8 mm, lățimea este de 0,31-0,33 mm. Este usor curbat, de forma cilindrica alungita, capatul opus locului de prindere este usor expandat. Pe
- În a 3-a zi, oul ia o poziție înclinată, iar în a 3-a zi, se află pe fundul celulei. La capătul liber expandat al oului există o gaură minusculă prin care spermatozoizii intră din recipientul uterin în timpul trecerii ovulului prin oviduct. Această gaură se numește micropil. Alteori, micropilul este impermeabil atât la bacterii, cât și la viruși. Oul are capete anterioare și posterioare. Capul larvei se dezvoltă în capătul anterior liber al oului.
Dezvoltarea embrionară începe cu fragmentarea nucleului. Nucleii fiice formați ca urmare a diviziunii sunt mai întâi localizați aleatoriu în interiorul gălbenușului, apoi se deplasează cu zone mici ale citoplasmei la periferia oului și formează un strat continuu de celule - blastodermul, care acoperă întreaga coajă de ou de la interiorul. Pe partea convexă a oului, celulele blastodermelor încep să crească și să se înmulțească mai repede decât celulele de pe partea concavă. Ca urmare, se formează o dâră embrionară sub forma unei șuvițe late. Acesta este începutul formării embrionului. Dâra germinativă crește datorită diviziunii și diferențierii celulare, formării țesuturilor. Ca urmare a acestor procese, are loc dezvoltarea larvei. În a doua jumătate a zilei a 2-a începe procesul de segmentare a embrionului. La capătul anterior al benzii germinale se formează un lob preoral (acron), pe care se formează un tubercul - rudimentul buzei superioare. Sub acest tubercul, apare o adâncitură - viitoarea gură. Apoi - rudimentele antenelor. Segmentele toracelui și abdomenului se desprind din segmentul capului înapoi. Din partea ventrală (ventrală) a acestor segmente apar excrescente - rudimentele membrelor pereche. Mai întâi apare pe piept, iar mai târziu pe abdomen. În același timp, în secțiunea capului se găsește o segmentare distinctă: se formează rudimentele a cinci segmente ale capului. În plus, apar găuri de respirație - stigmate, precum și începuturile glandelor de filare și vaselor malpighiene. Pe măsură ce se formează segmentele corpului și ale membrelor, are loc separarea părților individuale.
Ulterior, dâra germinativă crește în dimensiune. Reproducerea celulelor blastodermice duce la invaginarea părții mijlocii a benzii germinale (procesul de gastrulare), care coboară mai adânc în ou și se separă de blastoderm. În procesul de gastrulare, mai întâi se formează un șanț longitudinal pe suprafața benzii germinale, iar apoi marginile exterioare ale șanțului încep să crească unele spre altele. Ca urmare, viitorul al doilea strat germinativ se invaginează în interiorul benzii germinale. Când marginile șanțului se închid, se formează stratul exterior (ectoderm). Sub el se află stratul interior (mezodermul). La capetele stratului mezodermic apar invaginările, formate din grupuri de celule care formează al treilea strat germinativ (endoderm). Încep să apară rudimentele organelor individuale. Dezvoltarea și diferențierea lor continuă până când larva iese din celulă.
Din ectoderm se formează peretele corpului, intestinele anterior și posterior, sistemul traheal și nervos și multe glande; din endoderm - rudimentele înțepăturii, aripilor și membrelor, secțiunile de capăt ale tractului genital, sistemul nervos și organele senzoriale; din mezoderm - mușchi, grăsime, glande sexuale.
Un embrion complet format, care s-a transformat în esență într-o larvă, umple întregul ou. Ea începe să se miște intens, să atragă aer în trahee, să înghită lichid amniotic, crescând astfel volumul corpului ei. Larva este eliberată din membranele embrionare, rupe corionul (coaja secundară a oului, formată prin transformarea celulelor epiteliale foliculare) și eclozează, adică iese. De obicei, dezvoltarea embrionară a albinelor durează aproximativ 3 zile. Schimbările de temperatură pot crește sau scurta perioada de dezvoltare embrionară.
După eliberarea larvei din ou, începe dezvoltarea postembrionară, care este însoțită de metamorfoză (transformare).
Metamorfoza este o transformare profundă a structurii organismului, în urma căreia larva se transformă într-o insectă adultă. În funcție de natura dezvoltării postembrionare la insecte, se disting două tipuri de metamorfoză:
incomplet (hemimetabolism), când dezvoltarea unei insecte se caracterizează prin trecerea a doar trei etape - ouă, larve și faza adultă (adult);
complet (holometabolism), atunci când tranziția larvei la forma adultă se realizează într-un stadiu intermediar - stadiul de pupă.
Cu o transformare incompletă, modul de viață al stadiilor incipiente și al adulților este similar. Larva cu acest tip de dezvoltare este similară cu o insectă adultă și, ca și ea, are ochi compuși, organe bucale și apoi rudimentele aripilor care sunt clar vizibile din exterior. Rudimentele aripilor cresc în dimensiune cu fiecare năpârlire ulterioară, iar în ultima vârstă preimaginală pot acoperi deja mai multe segmente ale abdomenului. La o insectă din ultima vârstă preimaginală, structurile imaginare sunt pe deplin dezvoltate, iar un adult apare ca urmare a ultimei năpârliri complet formate. Transformarea incompletă este caracteristică ortopterelor, insectelor, homopterelor etc.
Cu metamorfoza completă, întregul ciclu de dezvoltare este caracterizat prin trecerea etapelor de ou, larvă, prepupă, pupă și adult (Tabelul 3).
- Durata etapelor de dezvoltare a indivizilor coloniei de albine, zile
Trecerea de la larvă la adult are loc în stadiul de pupă. Aceasta este o etapă nehrănitoare în care aparțin țesuturilor
În stadiile anterioare, se formează structuri imaginare. La majoritatea insectelor, acest stadiu este imobil. Pupa nu răspunde la stimuli externi și este îmbrăcată într-o acoperire groasă. La sfârșitul acestei etape, coaja explodează și apare o insectă adultă.
La insecte, reglarea metamorfozei este realizată de hormonii produși de glandele endocrine (glandele endocrine). În prezent, sunt cunoscuți trei hormoni de metamorfoză care controlează dezvoltarea postembrionară: hormonul juvenil, hormonul de napârlire (larva) și hormonul de activare. Hormonul juvenil este produs și stocat în corpurile adiacente, care sunt două structuri de celule mici care se află în spatele creierului, pe părțile laterale ale esofagului.
Dacă corpurile adiacente sunt îndepărtate dintr-o larvă tânără, atunci pupația va urma următoarea năpârlire, în ciuda faptului că, în starea sa normală, ar trebui să mai facă mai multe năpârliri. Acest hormon determină creșterea larvelor, controlează dezvoltarea unui număr de organe și procese. De exemplu, corpul gras nu crește și funcționează în absența acestui hormon.
Hormonul năpârlirii, sau ecdizona, este produs în glandele protoracice, care sunt situate în protoraxul larvei deasupra primului ganglion nervos lângă primul spiracol. Se sintetizează în corpul insectei din. colesterolul. Hormonul declanșează procesul de năpârlire și astfel reglează indirect creșterea și formarea structurii. Introducerea acestui hormon la insecte determină formarea de umflături pe cromozomi giganți. Analiza acestor umflături a arătat că în ele are loc un proces intens de formare a ARN. Se crede că primul rezultat al acțiunii ecdizonei este activarea genelor, apoi stimularea biosintezei ARN și formarea enzimelor corespunzătoare care asigură procesul de metamorfoză.
Hormonul activator este produs în celule neurosecretoare speciale, care sunt situate în partea dorsală anterioară a ganglionului supraesofagian. Aceste glande sunt mai bine dezvoltate la albinele lucrătoare, mai puțin la matci și chiar mai puțin la trântori. Hormonul de activare afectează reluarea activității insectelor după fiecare napârlire și stimulează formarea altor doi hormoni - ecdysone și juvenile.
Albina este o insectă cu metamorfoză completă (Fig. 33).
Stadiul larvar. Se caracterizează prin nutriție și creștere intensivă. Stadiul larvei de albine lucrătoare durează 6 zile, matca - 5, trântorul - 7. În primele zile, larvele albinelor primesc atât de multă hrană încât înoată la suprafața ei. Baza hranei este secretul hipofarinei.
| 1 g | 3 | 4 5 | 6 7 8 amperi; | 10 | 11 12 | 13 14 15 16 17 18 19 20 | 21 |
| w |
Zi
Orez. 33. Etapele dezvoltării albinelor
glandele geal ale albinelor doici - un lichid alb translucid. Laptele secretat de glandele albinelor doice este foarte hrănitor. Conține carbohidrați, proteine, grăsimi, minerale, vitamine din grupa B. Înainte de a da hrană larvei, albina lucrătoare, coborând capul în celulă, deschide și închide în mod repetat maxilarele superioare. Apoi atinge fundul celulei cu fălcile sale și eliberează alimente lichide. Alte albine-doice în același mod adaugă următoarele porții de lapte în celula cu larva. Pentru întreg stadiul larvar, există aproximativ 10 mii de vizite la o celulă cu o larvă.
Hrana larvelor tinere și a larvelor mai bătrâne nu este aceeași. În a doua perioadă de viață, larva albinei lucrătoare începe să primească terci - un amestec care este preparat de albinele doici din miere și polen.
În timpul etapei larvare, dimensiunile liniare ale larvelor de albine lucrătoare cresc de peste 10 ori. Astfel, lungimea unei larve proaspăt eclozate este de 1,6 mm, o larvă de 1 zi este de 2,6 mm, o larvă de 2 zile este de 6 mm, iar până la sfârșitul stadiului de larvă este de 17 mm (ocupă aproape complet). partea de jos a celulei). Lungimea larvei uterului ajunge la 26,5 mm până la sfârșitul etapei.
Nutriția intensivă oferă larvelor o creștere semnificativă a greutății corporale. Larva eclozată cântărește aproximativ 0,1 mg. Pentru primul
- zile, masa unei larve de albine lucrătoare crește de 45 de ori, iar masa unei larve de drone - de 85 de ori. Până la sfârșitul etapei, greutatea larvelor indivizilor muncitori crește de 1565 de ori, iar cea a mamelor de 2926 de ori.
În aparență, larva diferă puternic de albina adultă (Fig. 34). Are o formă asemănătoare unui vierme, coperți moi de culoare albicioasă. Corpul său este format dintr-un cap, un trunchi articulat și un lob anal.
Capul larvei este mic, obtuz-conic, baza conului este formată din capsula capului, care este împărțită printr-o sutură longitudinală.
Orez. 34. Structura externă (G) și internă (1G) a larvei de albine "\u003e
Orez. 34. Structura externă (D) și internă (1D) a larvei de albine:
A - vedere generală; B - vedere laterală a capului; C - pozitia larvei adulte la fundul celulei; G - vedere laterală; y - rudimente ale antenelor; vg - buza superioară; vh - maxilarul superior; opzh - deschiderea glandei de filare; ng - buza inferioară; p - gura; ndg - ganglion supraesofagian; PC - intestin anterior; a - aorta; sk - intestinul mijlociu; zht - corp gras; zpzh - germenul glandei sexuale; c - inima; zk - intestin posterior; ms - vas malpighian; bn - lanț nervos abdominal; pzh - fier de filare; pdg - ganglion subesofagian
în două părți convexe egale, așa-numiții obraji. Vârful conului din partea din față a capului este format din buza superioară și alte piese bucale. Partea capului, inclusiv buza inferioară, maxilarul superior și partea capsulei capului de care sunt atașate, este separată de capsula capului printr-un șanț adânc care trece pe fiecare parte între baza maxilarului superior. iar maxilarul şi se extinde până la marginile posterioare ale capului.
Mandibulele se extind de la capsula capului sub formă de organe în formă de con. Rudimentele antene sunt situate pe capsula capului deasupra bazei mandibulelor sub formă de tuberculi rotunzi.
Larvele de pe segmentele toracice nu au rudimente exterioare ale aripilor sau picioarelor. Nu există ochi compuși. Cu toate acestea, sub cuticula larvară sunt indicate rudimentele lor, care se numesc muguri imaginali. Mugurii imaginari ai picioarelor sunt fusiformi si disecati prin caneluri transversale.Rudimentele aripilor constau din doua procese plate, indoite spre interior ale hipodermului.Sunt situate pe laturile celui de-al doilea si al treilea segment toracic.In regiune din al optulea și al nouălea segment abdominal pe latura ventrală apar rudimentele organelor genitale.
Trebuie remarcat faptul că dezvoltarea aripilor, antenelor, ochilor și picioarelor nu are loc prin restructurarea structurilor larvare reduse; ele se dezvoltă din zone izolate ale țesuturilor embrionare care se află într-o stare inactivă, numite rudimente.
Larvele diferă de albina adultă prin structura organelor interne (vezi Fig. 34). Locul central în corpul larvei este ocupat de canalul intestinal, care constă din trei secțiuni - intestinul anterior, mijlociu și posterior. Intestinul anterior arată ca un tub scurt și subțire, ușor extins la capătul posterior. Intestinul anterior conține o cavitate bucală mică, un faringe scurt și esofag. Gușa de miere, caracteristică albinelor adulte, este absentă. În peretele intestinului anterior există mușchi care asigură ingestia de hrană lichidă. La joncțiunea esofagului și a intestinului mediu, există un mic pliu inelar care joacă rolul unei valve. Închide lumenul esofagului și împiedică conținutul intestinului mediu să curgă înapoi în esofag. Până la sfârșitul etapei larvare, în locul valvei se formează un sept gros, care acoperă intestinul mediu. 1
Intestinul mediu este cea mai mare secțiune a intestinului și ocupă cea mai mare parte a corpului larvei. O trăsătură distinctivă a acestei secțiuni este absența conexiunilor sale cu intestinul posterior, ca urmare a căreia părți nedigerate ale hranei rămân în ea pe tot parcursul etapei larvare. Legarea acestor două secțiuni are loc numai înainte de învârtirea coconului, când larva este deja sigilată. În această perioadă, membrana care separă intestinul mijlociu de intestinul posterior se sparge, iar fecalele trec în intestinul posterior, după care sunt îndepărtate din corp la fundul celulei.
În intestinul mijlociu, larvele au vase malpighiene. Albina are patru dintre ele. Sunt tuburi lungi, ușor întortocheate, care trec de-a lungul intestinului mediu. În timpul vieții larvare vasele malpighiene sunt închise. Abia până în a 6-a zi a etapei larvare, când se umflă puternic de la produsele de degradare acumulate acolo, capetele posterioare și conținutul se sparg, trec în intestinul posterior și apoi sunt îndepărtate. Astfel, larva nu își face nevoile în timpul creșterii și al hrănirii sale.
Intestinul posterior este mic și este un tub îngust, curbat la un unghi ascuțit și se termină în anus.
Inima larvei este formată din 12 camere cu aceeași structură (la o albină adultă sunt doar cinci) și este un tub cu pereți subțiri situat direct sub cuticula abdomenului și a pieptului. La al doilea segment al regiunii toracice trece aorta. Are aspectul unui tub, îndoit în jos, care trece apoi în cap și se termină la suprafața anterioară a creierului, unde hemolimfa se revarsă în cavitatea corpului. Pe suprafața aortei este o rețea densă de trahee.
Între toate camerele inimii există deschideri prin care hemolimfa pătrunde în inimă. Datorită faptului că capetele frontale subțiri ale fiecărei camere ies în camera următoare, nu este permisă curgerea inversă a fluidului.
Aparatul respirator al larvei este destul de simplu și este reprezentat de două trunchiuri traheale longitudinale mari care trec de-a lungul părților laterale ale corpului, cu mici ramuri din acestea care se răspândesc pe tot corpul. Pe părțile laterale ale segmentelor există 10 perechi de spiraculi, care sunt depresiuni înconjurate de inele chitinoase înguste.
Sistemul nervos este format din doi ganglioni ai capului aranjați simplu (un supraesofagian mare și unul subglot mic) și un cordon nervos ventral care se extinde de-a lungul părții inferioare a toracelui și a abdomenului. Lanțul abdominal este format din 11 ganglioni - 3 toracici și 8 abdominali. Ultimul ganglion abdominal se află la mijlocul celui de-al unsprezecelea segment; s-a format prin fuziunea ganglionilor ultimelor trei segmente. Ganglionii sunt conectați între ei prin conexiuni pereche. Ganglionul subesofagian este, de asemenea, conectat la cordonul nervos ventral prin două conjunctive.
Organele genitale sunt prezente în larva albinei lucrătoare în perioada incipientă și sunt reprezentate de două creste situate în al optulea segment. Lungimea lor este în medie de 0,27 mm, iar lățimea este de 0,14 mm. Pe parcursul întregului stadiu larvar are loc dezvoltarea primordiilor ovariene. La larva de 2 zile au dimensiuni nesemnificative, la larva de 6 zile sunt deja alungite și sunt situate spre capătul caudal prin al nouălea segment. Până la sfârșitul etapei larvare, numărul de tuburi de ou la o albină lucrătoare ajunge la 130-150. În etapele ulterioare, când larva se transformă în pupă, țesuturile larvare se dezintegrează și numărul tuburilor de ou la albina lucrătoare scade de la 20 la 3, în timp ce în uter, ovarele continuă să se dezvolte în stadiul de pupă.
Corpul gras este bine dezvoltat în larvă, unde are loc acumularea de nutrienți. La larvele mai în vârstă, corpul gras ajunge la 60% din greutatea corpului și umple aproape întreaga cavitate dintre intestinul mediu și cuticula larvei. Corpul gras este o concentrație de grăsimi, proteine și carbohidrați, datorită cărora pupa se dezvoltă în viitor. Corpul adipos, pe lângă celulele adipoase reale, este format și din celule excretoare și enocite care captează sărurile de acid uric.
Glanda de filare este prezentă numai în larve. Secreția acestei glande servește ca material pentru rotirea coconului înainte de tranziția larvei la stadiul de pupă. Glanda de filare arată ca două tuburi lungi, care sunt plasate sub intestinul mijlociu din partea ventrală. În zona anterioară, ambele tuburi sunt conectate într-un canal excretor nepereche, care se deschide sub orificiul bucal de la vârful buzei inferioare.
Pe parcursul întregului stadiu, larva dezvoltă rudimente de înțepăt, dintre care o pereche este situată pe al unsprezecelea segment și două perechi pe al doisprezecelea. Există un rudiment al deschiderii genitale între ele prin segmente. Rudimentul unei glande otrăvitoare mari apare abia spre sfârșitul stadiului larvar, iar la începutul stadiului de pupă se formează o mică glandă otrăvitoare din adâncirea învelișului pe al nouălea segment.
În a 5-6-a zi după eliberarea larvelor din ou, albinele încep să sigileze celula cu un capac de ceară. Pe lângă ceară, compoziția materialului care alcătuiește capacul include polen, apă și masă de hârtie. Capacul se dovedește a fi poros, datorită căruia aerul necesar respirației larvei și apoi pupei pătrunde în celulă. Din acest moment, larva își finalizează creșterea și încetează să se hrănească.
De îndată ce celula este sigilată, larva se îndreaptă în ea, pereții intestinali se contractă și reziduurile alimentare nedigerate, care au spart mai întâi peretele subțire al intestinului mijlociu, trec în cel gros și de acolo spre exterior. Larva depune excremente într-unul din colțurile celulei. După aceea, larva învârte un cocon, adică împletește suprafața interioară a celulei. Când învârte un cocon, larva folosește secreția glandei de filare, vasele malpighiene și secrețiile lipicioase de pe pereții corpului său. Procesul de filare a fost studiat și descris în detaliu de Velich (1930).
În timp ce învârte coconul, larva face mișcări rapide de tremur ritmic; pornind de la cap, unda se deplasează prin segmente. În 1 minut, larva face până la 280 de mișcări. Concomitent cu tremurul, larva face mișcări de balansare ale capului cu capătul anterior al corpului. Ele au natura unor curbe întrerupte, eliptice, neregulate de sus în jos sau de la dreapta la stânga. În același timp, o substanță transparentă curge din deschidere cu o supapă pe buza inferioară, pe care larva o secretă sub forma unui fir gros sau mai multor fire subțiri. În 2 zile, larva, întorcându-se, întărește pereții coconului. La rotirea unui cocon, comportamentul larvelor de albine lucrătoare, trântori și matci a fost similar, diferența fiind doar în forma coconului. Depinde de dimensiunea celulei în care se află larva. De exemplu, la o albină lucrătoare, coconul corespunde formei unei prisme hexagonale, al cărei fund este o piramidă cu trei laturi, iar capacul este o amprentă exactă a unei celule de albine sigilate. Coconul după ieșirea tinerei albine nu este îndepărtat. Datorită faptului că zeci de generații sunt afișate într-o singură celulă, diametrul celulei se îngustează în timp. Deci, dacă o celulă normală a unui pieptene proaspăt construit are un volum de 0,282 cm3 și o grosime de fund de 0,22 mm, atunci după 20 de generații de indivizi de albine, cubatura celulei scade la 0,248 cm3, iar fundul se îngroașă la 1,44 mm. .
Larva albinei lucrătoare și a matcii învârt un cocon timp de 2 zile, iar larva trântorului timp de 3 zile. Apoi ea năpădi a cincea oară. Această năpârlire este precedată de o serie de modificări în organismul larvei. Rudimentele tuturor anexelor situate sub cuticulă încep să se întoarcă pe dos sau să se deplaseze spre exterior și se transformă în părți externe ale corpului. Aceste organe iau treptat forma caracteristică pupei. Capul, piesele bucale și picioarele se rotesc dinspre anterior spre posterior-abdominal. După rotirea coconului, larva se îndreaptă și îngheață. Începe procesul pregătitor pentru stadiul de pupă, care necesită timp considerabil. Această perioadă se numește stadiul prepupal.
Stadiul prepupal. Durata etapei de prepupa pentru albina lucratoare este de 3 zile, pentru matca - 2, pentru trantor - 4 zile. Această perioadă este caracterizată de transformări semnificative în structura larvei. Cuticula larvară este complet separată de cuticula pupală sub influența unui hormon special de năparire ecdysone. O excepție o constituie conexiunile traheale dintre spiraculele larvare și pupale. Capul are deja 2/3 din dimensiunea capului adultului. Ochii sunt măriți, iar suprafața lor devine pliată și șifonată. Pieptul este separat de cap printr-o constricție clar vizibilă. Aripile încep să crească în lățime, în acest stadiu sunt încrețite și apăsate strâns pe piept. Părțile bucale se alungesc și se contopesc într-un grup compact.
Segmentele pieptului și abdomenului sunt împărțite de-a lungul unei linii orizontale în două jumătăți - dorsală și abdominală, dobândind treptat structura caracteristică unui adult.
În plus, organele interne sunt supuse cariilor. Acest proces se numește histoliză. Este însoțită de pătrunderea și introducerea în țesuturi a celulelor sanguine - fagocite. Sursele de energie necesare reacțiilor chimice care stau la baza acestor procese sunt rezervele de grăsimi, glicogen și alți carbohidrați din corpul gras al larvei, zahărul hemolimfei și mușchii. În timpul histolizei, hemocitele încep să funcționeze ca celule devoratoare, adică fagocitele, iar enzimele transformă corpul adipos, majoritatea mușchilor larvei, într-un substrat nutritiv furnizat de hemolimfă țesuturilor în creștere. În etapele ulterioare ale histolizei, fagocitele în sine sunt preluate de noile țesuturi în curs de dezvoltare. Sistemul muscular suferă cele mai mari modificări, în urma cărora prepupa este imobilizată.
O gușă de miere cu o valvă apare în intestinul anterior, intestinul mijlociu devine în formă de buclă și capătă o structură pliată. În intestinul posterior apar două secțiuni - intestinul subțire și rectul. Numărul de tubuli ai vaselor malpighiene crește. Se observă fuziunea parțială a ganglionilor lanțului neural, în urma căreia pupa are șapte ganglioni ai lanțului abdominal în loc de 11 ganglioni ai larvei. Corpul gras este redus în dimensiune.
Stadiul de prepupa se termină cu îndepărtarea vechii cuticule, iar pupa iese de sub ea.
Etapa de pupă. Întreaga suprafață a corpului pupei nou apărute este acoperită cu pliuri, dar nu șifonată. În structura sa, pupa este asemănătoare cu o albină adultă, deși, spre deosebire de aceasta din urmă, este nemișcată, corpul său este lipsit de pigmentare (alb). Cu toate acestea, procesele vieții interne nu încetinesc, ci decurg cu mare eficiență. Pupa imediat după terminarea ultimei năpârliri este acoperită cu o cuticulă. Stadiul pupal este caracterizat de transformări profunde. Sursa de substanțe plastice pentru formarea de noi organe și țesuturi, sau pentru histogeneză, sunt produsele de degradare transportate de hemocite. Glandele de filare, canalul digestiv, corpul adipos și mușchii se dezintegrează complet.
În loc de organe și țesuturi degradate ale larvei, se formează noi organe ale unei insecte adulte.
Capul este mult mai lung și extins aproape la dimensiunea capului adultului. Există o punte între capul extins și protoraxul îngustat. Cu toate acestea, este absent între segmentele toracice și abdominale. Pupa nu are încă aripi, dar antenele, proboscidele și picioarele sunt deja marcate clar.
Unele dintre elementele corpului gras în această perioadă sunt împrăștiate în cap și piept, iar unele sunt concentrate în abdomen. Odată cu vârsta, elementele corpului gras sunt mai uniform distribuite în cap, piept și abdomen. Structura stomacului se schimbă.
Inima se schimbă puțin: numărul de camere scade de la 12 la cinci. Pe tot parcursul etapei de pupă, inima nu încetează să funcționeze, deoarece procesul de metamorfoză necesită mișcarea hemolimfei. Masa ganglionului supraesofagian din cap crește. Acest lucru se întâmplă din cauza celulelor nervoase nediferențiate. În plus, ganglionii nervoși din piept se îmbină, precum și ultimii patru ganglioni de la capătul abdomenului.
Rudimentele gonadelor nu sunt distruse. Creșterea lor în stadiul inițial este asociată cu dezintegrarea celulelor corpului adipos și eliberarea de material plastic sub formă de albumide. Pe măsură ce se dezvoltă, vin în contact cu părți ale canalelor gonadale care decurg din ectoderm.
Unul dintre indicatorii vieții interioare a crisalidei este respirația acesteia.
Sistemul respirator suferă o serie de modificări în timpul histolizei și histogenezei. Deci, primul și al treilea spiracul toracic cresc, iar al doilea, dimpotrivă, scade. Distrus, trahee. Trahee noi, precum și sacii de aer, sunt formate din rudimente imaginare, care sunt situate pe trunchiurile traheale. 1
Ochii compuși cresc din mugurii de pe părțile laterale ale capului. După cum am menționat mai sus, pupa este inițial albă. În timpul dezvoltării în procesul de formare a diferitelor organe, culoarea tegumentului exterior al pupei se schimbă în următoarea secvență: la o zi după pupație, ochii compuși sunt încă alb pur, după 2 zile apare o nuanță gălbuie, pe A 14-a zi devin roz la culoare, iar în a 16-a zi - violet închis.
Articulațiile picioarelor sunt la început gălbui, din a 18-a zi articulațiile și ghearele de pe picioare capătă o culoare galben-maronie, călcâiele și părțile bucale sunt oarecum mai deschise. Cufărul devine fildeș. Din a 19-a zi, pieptul începe să se întunece ușor. Fildeș pe burtă. Capetele segmentelor picioarelor sunt maro. O întunecare mai semnificativă se observă la capătul bărbiei și la baza maxilarului superior. Ochii devin violet. În a 20-a zi, culoarea întregii pupe este gri închis.
Etapa de imago. La sfârșitul stadiului de pupă, pielea pupalei este aruncată, de sub care iese o albină complet formată. Cu mandibulele sale, albina roade prin capacul celulei și iese pe suprafața fagurelui. O albină proaspăt eclozată, în comparație cu o albină mai în vârstă, are un înveliș chitinos mai moale și, în plus, corpul ei este dens acoperit cu fire de păr. În timp, albinele își pierd o parte din linia părului, iar chitina devine mult mai dură. În timpul perioadei imaginare, nu apar modificări semnificative în structurile externe ale indivizilor de albine. "
Metabolismul în perioada postembrionară. Larva și pupa suferă procese metabolice intensive, în urma cărora se acumulează nutrienții necesari formării unei albine adulte.
Potrivit lui Strauss (1911), o caracteristică a metabolismului albinelor este acumularea unor cantități semnificative de glicogen ca energie pentru metamorfoză. Până la sfârșitul etapei larvare, conținutul de glicogen ajunge la 30% din masa uscată a larvei. În același timp, are loc o creștere a cantității de grăsime, creșterea celulelor adipoase.
În primul rând, glucoza este consumată în corpul albinelor. Conținutul său total în corpul larvelor de albine lucrătoare este de două ori mai mare decât în corpul larvelor mame. Când conținutul de glucoză din hemolimfă scade sub valoarea admisă, carbohidrații complecși încep să se descompună și să fie utilizați. Cea mai mare concentrație de glucoză se observă la mijlocul etapei deschise.
Trebuie remarcat faptul că activitatea majorității enzimelor studiate este cea mai mare la larvele tinere. Apoi scade și atinge un minim în stadiul de pupă. În același timp, dezvoltarea activității enzimatice la albinele lucrătoare este mai semnificativă decât la mătci. Există o opinie că în perioada de dezvoltare postembrionară, mătcile au un metabolism carbohidrați mai aerob în comparație cu indivizii muncitori, iar dezvoltarea accelerată a matcilor se explică prin activitatea mai mare a unui număr de enzime. Diferențele în cantitatea și calitatea mitocondriilor (organite celulare) la larvele uterine mai vechi decât
- vechi de o zi în comparație cu cele ale larvelor de albine lucrătoare de aceeași vârstă. În conformitate cu aceasta, unele diferențe sunt observate și în unele procese oxidative care au loc în mitocondrii.
La larva de matcă, curba consumului de oxigen crește până când celula este sigilată. Cu toate acestea, după etanșare, curba de absorbție a oxigenului scade (care coincide cu rotirea coconului) și atinge un minim în stadiul pupal inițial. Nivelul consumului de oxigen pe unitatea de masă la matci este cel mai scăzut în comparație cu albinele lucrătoare și cu trântorii.
Cantitatea de oxigen absorbită per individ în trântori în perioada postembrionară este mai mare decât la matci și albinele lucrătoare.
Coeficientul respirator, care caracterizează substratul folosit ca sursă de energie pe parcursul întregii etape de dezvoltare a larvei la o albină lucrătoare, se modifică, iar valoarea acestuia depășește unu. Dacă valoarea coeficientului respirator este mai mare de unu, atunci aceasta indică intensitatea proceselor asociate cu tranziția compușilor bogați în oxigen la compușii mai săraci în oxigen.
Creșterea coeficientului respirator în a doua perioadă de dezvoltare a larvelor coincide cu momentul trecerii la un tip de hrană calitativ diferit, în principal carbohidrați. După sigilarea celulei și până la pupația larvelor, coeficientul respirator este stabilit la 0,9.
Principalele surse de energie în larva și prepupa sigilate sunt carbohidrații depuși sub formă de glicogen, iar în a doua jumătate a stadiului de pupă, grăsimile.
În uter, spre deosebire de albina lucrătoare, coeficientul respirator scade până în a 2-a zi de dezvoltare larvară, ceea ce este caracteristic metabolismului grăsimilor. În a 4-5-a zi a aceleiași etape, coeficientul respirator este aproape egal cu valoarea coeficientului respirator al larvelor de albine lucrătoare, ceea ce indică transformarea carbohidraților în grăsime. În stadiul larvar sigilat, coeficientul respirator este sub unu, indicând descompunerea carbohidraților.
Din cele de mai sus, putem concluziona că schimbul de gaze la albinele este caracterizat de o serie de modele inerente insectelor. În timpul dezvoltării larvelor, intensitatea respirației, exprimată ca cantitatea de oxigen consumată pe unitatea de masă, scade. Cu toate acestea, cantitatea de oxigen consumată per larvă crește, atingând un maxim înainte de pupație. În stadiul de pupă, schimbul de gaze scade, crescând spre ieșirea adultului. În același timp, alături de asemănări, există diferențe în schimbul respirator al celor trei forme de albine.
La insecte, dezvoltarea este împărțită în două perioade - embrionară și postembrionară.
Dezvoltare embrionară. Celulele de ou de insecte sunt în cea mai mare parte de formă ovală și foarte bogate în gălbenuș, care umple toată partea centrală a oului. În consecință, zdrobirea oului este parțială, superficială; ca urmare, se formează un strat continuu de celule identice, blastodermul, care acoperă gălbenușul central. În același timp, o parte din nuclee rămâne în masa gălbenușului: în viitor, celulele gălbenușului speciale se formează pe cheltuiala lor. Pe viitoarea parte ventrală a embrionului, blastodermul se îngroașă. Această zonă se numește dâră germinativă; în viitor, pe cheltuiala sa, se formează partea principală a corpului embrionului.
În regiunea fâșiei germinale încep procesele de diferențiere. Ca urmare a reproducerii și pătrunderii în adâncurile celulelor blastodermice, se formează două benzi de celule - dungi mezodermice, iar blastodermul însuși devine ectodermul. Mai târziu, dungile mezodermice se împart în primordii pereche, dând naștere unui dublu rând de saci celomici. La insectele superioare (neg. Diptere) segmentarea dungilor mezodermice este suprimată și nu are loc separarea celomilor independenți.
Problema formării endodermului la insecte nu a fost încă pe deplin rezolvată. În multe forme, celulele viteline par a fi rudimentul endodermului. La Diptereși alte insecte, două grupuri de celule sunt separate la capetele anterioare și posterioare ale benzii germinative, care sunt considerate și începuturile endodermului.
Chiar de la începutul depunerii straturilor germinale începe procesul de formare a membranelor embrionare, care este extrem de caracteristic dezvoltării insectelor.
În cel mai simplu și mai răspândit caz, de-a lungul periferiei benzii germinale, ectodermul formează mai întâi o rolă, iar apoi un pliu în două straturi, crescând treptat din toate părțile până la banda germinativă, care în același timp este oarecum scufundată în galbenusul. În final, marginile pliului se închid peste rudimentul embrionului, lăsând între ele și acesta cavitatea amniotică ocupată de lichid, separată de mediul extern printr-o membrană dublă. Stratul exterior al membranei apare ca urmare a fuziunii straturilor exterioare ale pliului ectodermic care se inchide peste embrion si se numeste serosa, stratul interior din stratul interior al aceluiasi pliu, acesta din urma limitand direct cavitatea amniotica. și se numește amnios. Serosa și amnionul protejează embrionul de deteriorare.
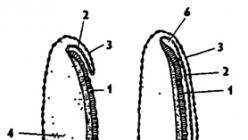
Între timp, dâra germinativă experimentează segmentarea, iar segmentele sunt izolate, ca și în celelalte Arthropodași inele, în ordine din față în spate. În secțiunea capului, sunt așezați lobii oftalmici și antenari cu rudimentele corespunzătoare, segmentul intercalar, care suferă ulterior reducerea și 3 segmente ale membrelor bucale. Mai posterior, se diferențiază 3 segmente toracice, purtând rudimentele membrelor, și 11 segmente abdominale.
În ceea ce privește diferențierea organelor interne, în principalele sale caracteristici constă în următoarele. Intestinul anterior și intestinul posterior sunt formate din două invaginări ectodermice. Problema originii intestinului mediu este mai complicată. Insectele se caracterizează printr-o varietate uimitoare de moduri și surse de formare a acestora din urmă. În unele cazuri, se formează din cauza celulelor endodermice de gălbenuș, în altele, elementele ectodermice iau parte la formarea intestinului mediu - celule speciale localizate la capetele închise orbește ale rudimentelor intestinului anterior și posterior. Și, în sfârșit, la o serie de specii (afidele femele partenogenetice, niște ihneumoni), intestinul mediu este format doar de ectoderm datorită creșterii puternice a anlagei intestinului anterior, care ajunge la capătul posterior al corpului. Elementele endodermice degenerează în același timp. Vasele malpighiene iau naștere din ectodermul intestinului posterior. Sistemul nervos este așezat sub forma unei creste ventrale a ectodermului, care apoi se scufundă sub tegument. Sacii celomici se dezintegrează, formând o cavitate corporală mixtă - un mixocoel. Elementele celulare eliberate în același timp dau naștere mușchilor, inimii, grăsimii corpului și altor organe. Celulele germinale primare sunt depuse foarte devreme, uneori chiar și în perioada în care gălbenușul este înconjurat de un singur strat de blastoderm.
Embrionul crește, gălbenușul hrănitor este consumat constant, iar membranele embrionare explodează, se înfășoară pe partea dorsală sub formă de pliuri, care suferă degenerare. Embrionul este gata să iasă din ou, iar cu aceasta se încheie dezvoltarea embrionară a insectei.
Dezvoltare postembrionară. La insecte, un animal tânăr sau o larvă cu un număr întreg de segmente iese din ou. Singurele excepții sunt criptomaxilarele primitive, aparținând neg. Protura, în care tinerii diferă de adulți prin absența a trei segmente abdominale posterioare. Acestea din urmă se formează din zona de creștere din spate, secvenţial după primele moarte. Astfel, dezvoltarea fără anamorfoză este tipică pentru insecte și doar în protur se găsește anamorfoză reziduală.
Dezvoltarea postembrionară a insectelor este caracterizată de o diversitate considerabilă, care este în mare măsură determinată de caracteristicile biologice ale reprezentanților acestui grup de animale.
În cele mai primitive forme (subcl. Entognatha) creșterea și dezvoltarea unui organism tânăr nu este însoțită de modificări structurale semnificative. Puieții în toate caracteristicile principale sunt similare cu organismul mamei, adică stadiul larvar și metamorfoza sunt absente. În esență, în acest caz avem de-a face cu dezvoltarea directă. Insectele înaripate, care sunt mai înalte din punct de vedere evolutiv, au, dimpotrivă, metamorfoză, iar după natura transformării, se împart în hemimetabolice - cu transformare incompletă și holometabolice - cu metamorfoză completă. Metamorfoza incompletă este caracteristică grupurilor inferioare organizate de insecte înaripate: ortoptere, gândaci, gândaci, eferice, libelule. Cu o transformare incompletă, larva care eclozează din ou în aspect general este deja în multe privințe similară cu organismul adult, deosebindu-se de acesta în principal prin starea embrionară a aripilor, subdezvoltarea caracteristicilor sexuale secundare și adesea prezența unor provizorii. , organe pur larvare: de exemplu, branhii traheale la larvele de efei etc. Aripile sunt la început doar sub forma unor rudimente exterioare abia vizibile. Netezirea diferenței dintre larvă și organismul adult are loc treptat, însoțită de un număr de nămolări.
Cu fiecare napârlire ulterioară, larva devine din ce în ce mai asemănătoare cu stadiul adult al insectelor, sau adulților (imago), rudimentele aripilor cresc etc.
La unele insecte primitive, numărul de năpârliri este foarte mare și poate ajunge la 25-30 (erice). Reprezentanții altor detașamente au mai puține linii (4-5).

La insectele holometabolice, dezvoltarea este diferită. Larva diferă foarte mult de animalul adult: nu are deloc rudimente exterioare ale aripilor, de multe ori are un număr diferit de membre etc. Caracterele larvare se păstrează în timpul unei serii de nămolări, adică pe toată perioada larvară a viaţă. Larvele insectelor holometabolice sunt extrem de diverse. Larva diferă de imago prin prezența unui număr de caracteristici primitive: o omonomie mai mare a disecției, absența aripilor, o dezvoltare slabă a organelor de simț (nu există ochi compuși, ci doar ocelli, antenele sunt foarte mici, etc.), și în multe cazuri și dispozitivul și numărul de membre. Foarte des, membrele gurii la vârsta larvară sunt aranjate după un singur tip (roșea în omizi de fluture), la adulți - diferit (suge piese bucale la fluturii adulți).
După cum s-a menționat deja, structura caracteristică larvei se păstrează pe toată perioada larvară: transformarea nu are loc treptat, dar modificările necesare metamorfozei sunt amânate până la momentul ultimei năpârliri larvare. În această perioadă au loc transformări atât de fundamentale ale organismului larvei încât continuarea funcțiilor fiziologice obișnuite, în special nutriția și mișcarea, devine dificilă, chiar imposibilă. Acest lucru se reflectă în faptul că larva, după ultima năpârlire, intră într-o stare de repaus - devine o crisalidă. Pupa constituie etapa cea mai caracteristică a transformării holometabolice. Cu cât starea de odihnă este mai completă, cu atât transformarea completă este mai bine exprimată.
Dezvoltarea individuală a unei insecte sau ontogeneza poate fi împărțită în 2 perioade - dezvoltare în interiorul oului, sau embrionară, și dezvoltare după părăsirea oului, sau postembrionară.
Majoritatea insectelor depun ouă. Un ou de insectă este o celulă care conține un nucleu, protoplasmă, deutoplasmă sau gălbenuș, necesar pentru nutriția și dezvoltarea embrionului. În plus, ouăle de insecte conțin adesea microorganisme simbiotice obținute de la mamă prin ovare (adică, transovarian). Oul este acoperit la exterior cu un corion - o coajă. Corionul are uneori o microsculptură distinctă, care servește ca semn de încredere pentru distingerea genurilor și chiar a speciilor de insecte în funcție de faza oului. Sub corion se află membrana adevărată sau gălbenușă a oului. Odată cu formarea corionului, micropilului se lasă o deschidere, care servește la trecerea spermatozoidului în timpul fertilizării. Oul are capete anterioare și posterioare care corespund poziției embrionului. În ovar, polul anterior al oului este îndreptat spre capul mamei, respectiv, se află părțile dorsale și ventrale ale oului.
Mărimea și aspectul ouălor sunt variate. Dimensiunea este de 0,02 - 0,03 mm pentru afide și trips, 8-10 mm pentru lăcuste. Ouăle sunt netede în afide, striate în linguri, albușuri, uneori echipate cu capac, acoperite cu riduri.
Forma de ou. Oval - gândaci, fluturi - alungit, semisferic - linguri, în formă de sticlă - fluturi diurni, în formă de butoi - gândaci, au o tulpină - aripioare, ventuze. Ovipozitia: individual, in grupuri, deschis sau in substrat, poate fi protejata, acoperita cu secretii gonadale.
Dezvoltarea embrionară începe cu fragmentarea nucleului și deplasarea nucleelor fiice rezultate cu zone mici de protoplasmă către periferia oului. Aici, din masa nucleelor fiice, se formează un strat continuu de celule - blastodermul. Ulterior, blastodermul se diferențiază în zonele germinale și extra-embrionare: celulele acestuia din urmă nu participă la formarea embrionului, în timp ce celulele zonei germinale încep să se dividă mai intens și formează o bandă embrionară pe partea ventrală. a oului, care apoi iese și iau naștere ectodermul și mezodermul.
Dezvoltarea embrionului este însoțită de blastokineză: se formează membrane embrionare, se separă segmentele capului, toracice și abdominale. Bazele structurii interne încep să fie puse odată cu formarea mezodermului. Din ectoderm provin toate învelișurile exterioare - apoi gura și anusul, intestinele anterior și posterior. Mezodermul dă naștere sistemului muscular, corp gras, vas dorsal, teaca glandelor sexuale.
Embrionul începe să facă mișcări, captează aer, lichid din ou, corionul se rupe și embrionul, care s-a transformat în larvă, iese.
2. Reproducerea insectelor are o serie de caracteristici care constituie un aspect esențial al biologiei lor și necesită luarea în considerare a acestora. Aceste caracteristici includ metode de reproducere, nutriție suplimentară, întâlnirea sexelor și fertilizare, productivitatea sexuală.
Metode de reproducere .
Reproducerea la majoritatea insectelor este însoțită de împerechere și fertilizare, adică. asociat cu participarea a două sexe - prin urmare se numește gamogenetic bisexual (bisexual). Majoritatea insectelor depun ouă, dar există și alte metode de reproducere: naștere vie, partenogeneză, pedogeneză, poliembrionare. Nașterea vie este că dezvoltarea embrionară este finalizată în corpul mamei, prin urmare, în loc să depună ouă, se nasc în lume larve (afidele, tafanul, muștele suge de sânge). Larvele eclozează în tuburile de ou sau în timp ce trec prin oviducte.
Partenogeneza - reproducerea virgină se caracterizează prin absența fertilizării și se observă atât la insectele ovipare, cât și la cele vivipare.
Forme de partenogeneză:
1. arrhenotokia- numai masculii se dezvolta din oua nefertilizate; 2) thelytokia - numai femelele se dezvoltă din ouă nefertilizate; 3) amfitochie- masculii si femelele se dezvolta din oua nefertilizate.
Partenogeneza poate fi facultativă, permanentă și ciclică.
Partenogeneza facultativă sau spadică se manifestă inconsecvent, apare sub influenţe externe sau la o anumită stare fiziologică a femelei ouătoare (molia ţigănească, molia şoimului plopului).
Partenogeneza constantă de tip arrhenotoky este tipică insectelor sociale. La albine, ouăle nefertilizate se dezvoltă întotdeauna în masculi, iar din ouăle fecundate, femele. Partenogeneza permanentă de tip thelytoky se observă la insectele care nu au un mod de viață colonial. Deci, la mulți viermi, trips, muște și unii călăreți, masculii sunt absenți, reproducerea are loc întotdeauna prin partenogeneză și doar femelele eclozează din ouă nefertilizate.
Partenogeneza joacă un rol foarte important în viața insectelor. Datorită partenogenezei și modificărilor acesteia, potențialul de reproducere al speciei crește dramatic: odată cu reproducerea partenogenetică, șansele speciei de a supraviețui în condiții nefavorabile și în timpul așezării sunt semnificativ crescute.
3. După părăsirea oului, începe dezvoltarea postembrionară a insectei. Această perioadă nu este o simplă creștere și creștere a dimensiunii corpului, ci se caracterizează prin trecerea corpului de la o fază la alta. Acest tip de dezvoltare individuală se numește metamorfoză sau dezvoltare cu transformare.
Esența metamorfozei constă în faptul că un individ în curs de dezvoltare suferă o restructurare semnificativă a organizării sale morfologice și a caracteristicilor biologiei în timpul vieții sale. În acest sens, există o diferențiere a dezvoltării postembrionare în 2 faze - larvară și adultă. În faza larvară are loc creșterea și dezvoltarea indivizilor, în faza imago, reproducerea și așezarea. În alte cazuri, între aceste faze apare o fază intermediară - pupa. Există 2 tipuri principale de metamorfoză - transformarea incompletă și completă. conversie incompletă sau hemimetamorfoză 3 faze - ou, larvă, adult. Larvele sunt asemănătoare cu adulții (ochi compuși, piese bucale, rudimente ale aripilor, un mod de viață similar), asemănătoare imago. Cu complete (holometamorfoză) 4 faze. Larvele nu arată deloc ca adulții, nu există ochi, aripi complexe, trăiesc în condiții diferite.
Unii au picioare ventrale, un aparat bucal diferit, alții au glande de mătase sau de păianjen.
Modificări ale transformării incomplete: a) hipomorfoza - o transformare incompletă simplificată și este caracteristică insectelor înaripate care și-au pierdut aripile (păduchii secundari fără aripi, păduchii, reprezentanți fără aripi ai lăcustelor, lăcustelor, greierii, gândacilor, insectelor stick, mâncătorilor de fân, ploșnițelor); b) hipermorfoza - un ciclu incomplet complicat (aleurodide, trips, masculi coccizi). Apariția la sfârșitul fazei a larvelor din starea de repaus.
Anamorfoza- larvele sunt asemănătoare cu adulții dar au un număr mai mic de segmente abdominale; pe măsură ce larva se dezvoltă, segmente suplimentare cresc în partea de sus a abdomenului, dar numărul complet ajunge doar în faza adultă.
protomorfoza– năpârlirea în stare adultă, o oarecare asemănare a larvei cu faza adultă, dar absența divizării larvei în torace și abdomen (podura, cu două cozi, cu peri).
hipermetamorfoză- complicație a transformării complete (prezența a 2 forme de larve, iar uneori pupe (vezicule)).
4. Faza larvară este imediat după ecloziune din ou. Inițial ușoară, după ecloziune larva este incoloră, are coperți moi. La insectele vii în mod deschis, larva se întunecă și se întărește rapid. Stimulii de hrănire se creează după digestia rămășițelor gălbenușului embrionar și excreția excrementelor. Larva intră într-o pereche de nutriție, creștere și dezvoltare îmbunătățite. Creșterea și dezvoltarea lor sunt însoțite de năpârlirea periodică, vărsarea cuticulei pielii - datorită nămolării, corpul crește și apar modificări majore. Numărul de legături variază 3 (muște), 4-5 (bugs), 25-30 (padenki).
După fiecare năpârlire, larva intră în următoarea etapă sau în vârstă, prin urmare, năpârlirile împart vârsta larvelor. În concordanță cu numărul de nămolări, se găsește și numărul de stadii larvare. Stabilirea vârstei de năpârlire (necesară pentru planificarea momentului luptei) se realizează în funcție de următoarele caracteristici: antene la lăcuste, la creșterea dimensiunii corpului (lățimea capsulei capului) la omizi de fluturi și larve de gândaci. Larvele sunt de 2 tipuri - adulte (primare) și secundare.
Naiadele - larve cu branhii, trăiesc în apă (libelule, padyonki, muște de pietre).
Campodeoid - mobil, de culoare închisă, cu tegument dens și cu 3 perechi de picioare pectorale, un cap prognatic bine izolat (gândaci de pământ, înotători, ramuri).
Asemănător viermilor - sedentar, de culoare deschisă, lipsit de picioare abdominale.
Omizile au pielea capului bine despărțită, 3 perechi de picioare toracice, 2-8 perechi de abdominale.
faza pupală. Această fază de dezvoltare a metamorfozei este caracteristică numai insectelor cu ciclu complet. O caracteristică a pupei este incapacitatea de a se hrăni și de foarte multe ori a rămâne imobil. Trăiește din rezervele acumulate de larvă și este adesea privită ca o fază latentă.
În exterior, pupa nu arată ca un adult, dar are o serie de semne ale fazei adulte: rudimente externe ale aripilor, picioarelor, antenelor, ochilor compuși etc. Histoliza și histogeneza apar în faza pupală.
Histoliza- dezintegrarea organelor interne ale larvei, care este însoțită de pătrunderea și pătrunderea în țesuturile celulelor sanguine - hemocite. Hemocitele funcționează ca mâncători de celule, adică. fagocite, a căror activitate crescută duce la distrugerea și absorbția materiei tisulare. Histoliza captează sistemul muscular, astfel încât prepupa devine imobilă, afectează sistemul digestiv, dar nu afectează sistemul nervos și reproductiv, precum și vasul dorsal.
Histogenie- procesul de creare a țesuturilor și organelor vieții imaginare. Sursa pentru formarea acestor noi țesuturi și organe sunt produsele histolizei. Histogenia cuprinde sistemele musculare și digestive, rearanjandu-le pentru funcții noi, imaginare. Sistemul muscular este reconstruit pentru noi forme de mișcare pentru zbor, sistemul digestiv pentru noi tipuri de alimente.
În timpul histogenezei, rolul principal îl au rudimentele imaginare - grupuri de celule hipodermice din care provin anumite țesuturi și organe.
Hormonul năpârlirii este exidonă, care este necesară pentru dezvoltarea normală a larvelor.
Există următoarele tipuri de pupe:
pupe deschise- libere, presate doar pe corp anexele imaginare (antene, picioare, aripi), caracteristice gândacilor.
Pupele cu mandibule mobile - folosesc fălcile superioare mobile pentru a ieși din cocon și se pot mișca ele însele (reticulate, caddisflies, molii dințate). Pupele cu mandibule imobile nu le pot folosi pe acestea din urmă când ies din cocon (gândaci, himenoptere, fanoptere, multe diptere).
Păpuși ascunse acoperit cu piele de larvă întărită, care nu revarsă, care joacă rolul de cochilie sau de cocon fals - pupari (muște).
acoperit- au anexele imaginare strâns presate și lipite de corp datorită faptului că în timpul ultimei naparliri larva secretă un secret care, întărit, acoperă pupa cu o coajă tare (fluturi, gărgărițe).
Înainte de pupație, unele larve se înconjoară cu un cocon (mătase, pânză de păianjen). Uneori, locul pupației este tulpini de plante, leagăn de pământ, pupație deschisă.
Faza de imagine.
Insectele care ies din pupă au semne de adult, dar la început aripile rămân îndoite. Apoi își întind aripile, tegumentul se îngroașă și capătă culoarea unei insecte adulte complet formate. În faza adultă, insectele nu năparesc și nu sunt capabile să crească. Excepțiile sunt padyonki, proști. Funcția biologică a fazei adulte este de a se dispersa și de a se reproduce. Aceasta este deja o funcție a vieții speciei și are ca scop menținerea existenței speciei. Datorită aripilor, capacitatea insectelor adulte de a se așeza crește foarte mult, iar reproducerea vă permite să lăsați descendenții într-un loc nou.
Răspândirea insectelor adulte are loc atât prin zbor activ, cât și pasiv. Zborurile active sunt de obicei caracteristice insectelor mari și sunt observate la o serie de specii de libelule, lăcuste, fluturi, gândaci și sunt larg răspândite. Zborurile pasive sunt caracteristice afidelor și muștelor.
Trecerea la adulți este însoțită de modificări externe ale culorii corpului, o creștere a dimensiunii abdomenului la femelă, datorită dezvoltării ovarelor umplute cu ouă - la femelele fertilizate de termite și furnici - căderea aripilor, lăcustele imature - roz. , matur - galben strălucitor.
Diferențele de dimorfism sexual între bărbați și femele într-o serie de caracteristici sexuale externe, secundare - în forma și dimensiunea antenelor, dimensiunea corpului, diverse detalii structurale (gândacul rinocerului, gândacul căpriorului). Bărbații sunt mai mobili, duc un stil de viață mai deschis.
Polimorfism - existența insectelor, forme diferite în exterior ale aceleiași specii - (furnici, albine, termite, masculi, femele, muncitori, soldați). Polimorfismul sexual este controlat în cadrul familiei și nu depinde de influența factorilor externi.
Polimorfismul ecologic apare sub influența mediului extern (gangăci cu aripi lungi, cu aripi scurte, fără aripi).