Die Embryonalentwicklung von Insekten beginnt typischerweise mit der oberflächlichen Zerkleinerung des Eies, was mit dem reichen Gehalt des letzten nährstoffreichen Eigelbs verbunden ist. Das auf der Oberfläche des Eies gebildete Bastoderma ist an einigen Stellen der Oberfläche dicker; Diese Verdickung führt zum Keimstreifen und definiert die ventrale Seite des Embryos.
Die am Rand des Keimstreifens gebildete Ringfalte, die oberhalb des Keimstreifens überwuchert und zusammenwächst, ergibt zwei Schichten von Keimhäuten – die innere (Amnion) und die äußere (Serosa). Von der Zellschicht des Keimstreifens auf der dem Dotter zugewandten Seite,das primäre Endoderm wird abgetrennt – der gemeinsame Keim von Endoderm und Mesoderm. Anschließend wird der Keimstreifen segmentiert.
Die postembryonale Entwicklung von Insekten ist aufgrund der Vielfalt der Existenzbedingungen jugendlicher Insekten und des Grades ihrer Anpassung an diese sehr vielfältig. Es können zwei Haupttypen der postembryonalen Entwicklung unterschieden werden: unvollständige Transformation, Hemimetabolismus, und vollständige Transformation, Holometabolismus. Der Holometabolismus ist durch tiefgreifende Veränderungen bei Jungtieren gekennzeichnet, wodurch sich die Jungtiere in ihrer Struktur stark vom erwachsenen Insekt (Erwachsenen) unterscheiden. Daher erfordert der Übergang vom juvenilen Zustand, also von der Larvenphase zur Imaginalphase, eine radikale Umstrukturierung der Organe; Dies wird durch die Teilnahme einer Zwischenphase zwischen der Larven- und der Imaginalphase erreicht, in der die Jungtiere ruhen. Diese Zwischenphase ist die Puppenphase. Während des Puppenstadiums werden die inneren Organe des Insekts entweder mehr oder weniger umstrukturiert oder vollständig zerstört (Histolise) und neu aufgebaut; Im letzteren Fall erfolgt die Neubildung des Organs durch die sogenannten Imaginalscheiben, die aus Zellen embryonaler Natur (Histoblasten) bestehen. Somit ist die Puppenperiode eine Periode der aktiven Histolyse und Histogenese (Gewebeneoplasmen). Äußerlich repräsentiert die Puppe die Imaginalphase mit ihren charakteristischen Organen, die jedoch unterentwickelt blieben.
Die Entwicklung höherer Insektenordnungen erfolgt holometabolisch bei Käfern, Schmetterlingen, Köcherfliegen, Flöhen, Hautflüglern und Zweiflüglern.
Im Falle einer unvollständigen Transformation - Hemimetabolismus - gibt es keinen so tiefgreifenden Unterschied in der Struktur der jungen und erwachsenen Phase: Die jugendliche Phase ähnelt in ihrer Struktur der imaginären Phase mehr, dann etwas weniger. Schließlich in Fällen, in denen der Übergang von der jungen Phase erfolgt Phase bis zum Erwachsenen beschränkt sich nur auf das Wachstum und die Ablösung (im Zusammenhang mit dem Wachstum) der alten Hülle (Exuvia), man spricht von Amöbolismus, also dem Fehlen einer Transformation; Ein Beispiel für Amöbolismus ist die Entwicklung von Collembola bei den Apterygota. Allerdings kommt es meist bei aufeinanderfolgenden Stadien, die durch Häutungen, also durch das Abwerfen der Hülle des Voralters, voneinander getrennt werden, auch zu Veränderungen in der Form der Organe: zum Beispiel der Länge der Fühler und Cerci nimmt relativ zu, die Anzahl der Segmente nimmt zu; Einige Organe können wieder zum Vorschein kommen, beispielsweise die äußeren Genitalanhangsgebilde. Dies ist die Entwicklung bei einigen Apterygota, wie Thysanura und Diplura, und bei den meisten hemimetabolischen Pterygota. Bei einigen Pterygota, die sich mit Hemimetabolismus entwickeln (Libellen, Eintagsfliegen, Steinfliegen und Teile von Arthropoden), ist die Entwicklung dadurch erschwert, dass Jungtiere unter anderen Bedingungen leben als Erwachsene; So entwickeln sich Jungtiere von Libellen, Eintagsfliegen und Steinfliegen in Gewässern und erwerben in diesem Zusammenhang Organe der Wasseratmung – Kiemen, die ein Beispiel für larvale (provisorische) Organe sind. Die Existenzbedingungen spiegeln sich auch in der Entwicklung anderer Organe der Larve wider, die sich an die Umwelt anpassen, beispielsweise in der Umwandlung der Vorderbeine der Jungzikade in ein Graborgan im Zusammenhang mit ihrem unterirdischen Leben.
Insektenentwicklung
Die individuelle Entwicklung von Insekten (Ontogenese) besteht aus der Embryonalentwicklung in der Eiphase und der postembryonalen Entwicklung – nachdem die Larve das Ei verlassen hat, bis zum Erreichen der Erwachsenenphase – der Imago.
Embryonale Entwicklung. Insekteneier variieren aufgrund der Anpassung an die Umgebung, in der sie sich entwickeln, in ihrer Form. Bei Käfern beispielsweise sind die Eier überwiegend oval und entwickeln sich häufiger in einem geschlossenen Substrat; bei Wanzen - tonnenförmig, am Untergrund befestigt; Schmetterlinge – turmförmig oder flaschenförmig; bei Florfliegen (goldene Augen) Eier mit Stiel. Eier werden oft in Gruppen gelegt. Eiergelege können offen oder geschlossen sein. Ein Beispiel für offenes Legen sind die Eier des Kartoffelkäfers, die vom Weibchen an die Unterseite von Kartoffelblättern geklebt werden. Geschlossene Gelege umfassen Eikapseln aus Akridoiden, die aus Bodenpartikeln gebildet werden, die durch Ausscheidungen der Adnexdrüsen von Weibchen zementiert werden. Kakerlaken legen ihre Eier in Ootheken ab – Eikapseln, die sich im weiblichen Genitaltrakt bilden.
Insekteneier sind außen mit einer Schale – Chorion – bedeckt, die sie vor dem Austrocknen schützt (Abb. 337). Auf der Oberfläche der Schale befindet sich eine Mikropyle – ein kleines Loch mit einem komplexen „Pfropfen“ mit einem Röhrchen im Inneren für das Eindringen von Spermatozoen während der Befruchtung. Unter dem Chorion befindet sich eine dünne Dottermembran und darunter eine dichte Zytoplasmaschicht. Der zentrale Teil des Zytoplasmas ist mit Eigelb gefüllt. Das Zytoplasma enthält den Zellkern und die Polkörperchen.
Zerquetschen ist oberflächlich. Zunächst teilt sich der Kern viele Male, die Tochterkerne mit Abschnitten des Zytoplasmas wandern an die Peripherie des Eies, werden mit einer Membran bedeckt und es bildet sich eine Oberflächenschicht aus Zellen – das Blastoderm, und das Eigelb verbleibt in der Mitte des Eies Ei. Auf der ventralen Oberfläche des Blastoderms liegen die Zellen höher und bilden eine Verdickung – den Keimstreifen. Dieses Stadium der Embryonalentwicklung von Insekten entspricht der Blastula.
Die Zellteilung des Keimstreifens führt zur Entwicklung des Embryos. Der Keimstreifen sinkt allmählich ab und bildet die Bauchfurche.
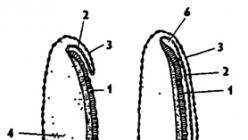
Reis. 337. Die Struktur eines Insekteneis (von Bei-Bienko): 1 - Mikropyle, 2 - Chorion, 3 - Dottermembran, 4 - Kern, 5 - Polkörperchen, 6 - Dotter

Die Falten des Blastoderms oberhalb der Furche schließen sich und es bilden sich Keimhäute: Serosa und Amnion (Abb. 338). Hier zeigt sich eine Konvergenz mit höheren Wirbeltieren, die ebenfalls über ähnliche Schalen verfügen. Aufgrund der gebildeten Fruchthöhle befindet sich der Embryo im Inneren der Eizelle in einem Schwebezustand, der ihn zuverlässig vor mechanischer Beschädigung schützt. Darüber hinaus erleichtert die Flüssigkeit, die die Fruchthöhle füllt, die Stoffwechselprozesse des Embryos.
Der Keimstreifen differenziert sich weiter in zwei Schichten: das untere Ektoderm und das obere Endomesoderm. Entomesoderm kann bei verschiedenen Insektenarten auf unterschiedliche Weise gebildet werden: durch Einstülpung oder Zelleinwanderung.
Im nächsten Entwicklungsstadium beginnt sich die ektodermale Schicht des Streifens an den Seiten nach oben zu biegen und schließt sich dann auf der Rückseite, wodurch eine geschlossene Wand des Embryos entsteht. Wenn sich die Körperwände auf der Rückseite schließen, gelangt ein Teil des Dotters und der Dotterzellen in den Körper des Embryos. Gleichzeitig mit der Bildung der Körperwände des Embryos werden im Entomesoderm am vorderen und hinteren Ende des Körpers zwei Zellgruppen getrennt. Dies sind die beiden Rudimente des Mitteldarms. Anschließend beginnen sich aus diesen beiden Primordien der vordere und hintere Abschnitt des Mitteldarms zu bilden, die dann verschmelzen. Gleichzeitig bilden sich am vorderen und hinteren Ende des Körpers des Embryos tiefe Einstülpungen des Ektoderms, aus denen sich der vordere und hintere Darmabschnitt bildet. Dann werden alle drei Abschnitte miteinander verbunden und bilden einen durchgehenden Darmschlauch.
Der mesodermale Streifen zerfällt in paarige metamere Rudimente von Zölomsäcken. Doch später zerfallen sie, und aus dem Mesoderm entstehen die Muskeln des Embryos, die somatische Schicht des Zölomepithels, das Herz, der Fettkörper und die Gonaden. Die viszerale Schicht des Zölomepithels wird bei Insekten nicht gebildet und die Körperhöhle wird durchmischt – das Mixocoel. Zölomrudimente verschmelzen mit der primären Körperhöhle.
Später werden aus dem Ektoderm das Nervensystem und das Trachealsystem gebildet. Malpighische Gefäße bilden sich aus den Wänden des hinteren Darms.
Im Laufe der Entwicklung durchläuft der Insektenembryo eine Segmentierung, die sich zunächst im vorderen Teil und dann im hinteren Teil des Körpers manifestiert. Im Kopfteil ein Akron mit Auge, Labial und

Antennenlappen, ein Interkalarsegment und drei Kiefersegmente. Dann werden drei Brust- und zehn Bauchsegmente sowie ein Anallappen gebildet.
Bei vielen Insekten durchläuft der Embryo drei Stadien, die durch eine unterschiedliche Zusammensetzung der Gliedmaßenprimordien gekennzeichnet sind: Protopod, Polypod und Oligopod (Abb. 339).
Die Embryonalentwicklung von Insekten ist durch das Phänomen der Blastokinese gekennzeichnet. Dabei handelt es sich um eine Positionsänderung des Embryokörpers im Ei, bei der die Eigelbreserven am meisten ausgenutzt werden.
Zwei Arten der Blastokinese bei Insekten werden von A. G. Sharov beschrieben. Bei Insekten mit unvollständiger Transformation befindet sich der Embryo zunächst mit dem Rücken nach oben und dem Kopf zum vorderen Ende des Eies, und dann, wenn sich die Fruchthöhle bildet, dreht sich der Embryo mit der Bauchseite nach oben und dem Kopf um. befindet sich jeweils auf der Rückseite des Eies.
Die Blastokinese verläuft bei den meisten Insekten unterschiedlich mit vollständiger Transformation und bei Orthopteren, bei denen der Embryo im Eigelb versinkt, ohne die Position des Körpers im Ei zu verändern.
In der Embryonalentwicklung von Insekten manifestieren sich Anpassungen an das Leben an Land: Schutzmembranen (Chorion, Serosa, Amnion), ein Nährstoffvorrat (viel Eigelb), eine mit Flüssigkeit gefüllte Fruchthöhle.
Vor dem Schlüpfen schluckt die gebildete Insektenlarve Flüssigkeit aus der Fruchthöhle, wodurch der Körperturgor zunimmt. Die Larve durchbricht mit ihrem Kopf, der oft Eierzähne oder einen Dorn trägt, das Chorion.
Postembryonale Entwicklung. Während der postembryonalen Entwicklung von Insekten, nach dem Schlüpfen aus dem Ei, erfolgt das Wachstum eines jungen Tieres durch aufeinanderfolgende Häutungen und den Durchgang qualitativ unterschiedlicher Entwicklungsphasen. Während der Ontogenese oder individuellen Entwicklung häuten sich Insekten 3-4 bis 30 Mal. Im Durchschnitt beträgt die Anzahl der Links 5-6. Der Zeitraum zwischen den Häutungen wird als Stadium bezeichnet, der Entwicklungsstand als Alter. Morphologische Veränderungen während der Entwicklung von der Larve zum erwachsenen Tier werden als Metamorphose bezeichnet. Bei allen Insekten, mit Ausnahme der unteren flügellosen Formen, hören Wachstum und Häutung der Erwachsenen nach Erreichen des Erwachsenenstadiums auf. Daher können beispielsweise Unterschiede in der Größe von Käfern derselben Art nicht auf verschiedene Altersgruppen zurückgeführt werden, sondern sollten lediglich als Ausdruck individueller Variabilität betrachtet werden.
Es gibt drei Haupttypen der postembryonalen Entwicklung von Insekten: 1) direkte Entwicklung ohne Metamorphose – Ametabolismus oder Protometabolismus; 2) Entwicklung mit unvollständiger Transformation oder mit allmählicher Metamorphose, - Hemimetabolismus; 3) Entwicklung mit vollständiger Transformation, d.h. mit ausgeprägter Metamorphose - Holometabolismus.
Ametabolismus, oder direkte Entwicklung, wird nur bei primären flügellosen Insekten aus der Borstenschwanzordnung (Thysanura) beobachtet, zu denen auch der Silberfisch (Lepisma) gehört. Die gleiche Art der Entwicklung wird bei Entognatha beobachtet: Springschwanz (Collembola) und Zweischwanz (Diplura).
Beim Ametabolismus schlüpft aus dem Ei eine Larve, ähnlich einem Erwachsenen. Die Unterschiede betreffen lediglich die Größe, die Körperproportionen und den Entwicklungsgrad der Keimdrüsen. Im Gegensatz zu geflügelten Insekten häuten sie sich auch im imaginären Zustand weiter.
Hemimetabolismus- unvollständige Transformation oder Entwicklung mit allmählicher Metamorphose. Es ist charakteristisch für viele geflügelte Insekten, zum Beispiel Kakerlaken, Heuschrecken, Heuschrecken, Wanzen, Zikaden usw.
Beim Hemimetabolismus schlüpft aus dem Ei eine Larve, ähnlich wie bei Erwachsenen, jedoch mit rudimentären Flügeln und unterentwickelten Keimdrüsen. Solche adulten Larven mit Flügelansätzen werden Nymphen genannt. Dieser Name ist der antiken griechischen Mythologie entlehnt und bezieht sich auf göttliche geflügelte Wesen in Form von Mädchen. Insektennymphen häuten sich mehrmals und mit jeder Häutung nehmen ihre Flügelrudimente zu. Eine ältere Nymphe häutet sich und aus ihr schlüpft eine geflügelte Imago. Abbildung 340 zeigt die Phasen der Heuschreckenentwicklung (Eier, Nymphen des 1.-5. Larvenstadiums und adulte Tiere) als Beispiel für eine unvollständige Transformation. Diese typische unvollständige Transformation wird Hemimetamorphose genannt.
Bei Insekten mit unvollständiger Metamorphose gibt es Entwicklungsfälle, bei denen sich Nymphen aufgrund spezieller Larvenanpassungen – provisorischer Organe – merklich von Erwachsenen unterscheiden. Eine solche Entwicklung wird bei Libellen, Eintagsfliegen und Steinfliegen beobachtet. Die Nymphen dieser Insekten leben im Wasser und werden daher so genannt

Reis. 340. Entwicklung mit unvollständiger Metamorphose bei der Heuschrecke Locusta migratoria (nach Kholodkovsky): 1 - Prothorax, 2 - Mesothorax mit Flügelknospen, 3 - Metathorax mit Flügelknospen
Najaden (Wassernymphen). Sie haben provisorische Organe wie Trachealkiemen, die bei erwachsenen Landtieren verschwinden. Und Libellenlarven haben immer noch eine „Maske“ – eine modifizierte Unterlippe, die zum Greifen von Beute dient.
Holometabolismus- vollständige Transformation. Die Entwicklungsphasen in der Holometabolie sind: Ei – Larve – Puppe – Imago (Abb. 341). Diese Entwicklung ist typisch für Käfer, Schmetterlinge, Zweiflügler, Hautflügler, Köcherfliegen und Florfliegen.

Reis. 341. Entwicklung mit vollständiger Transformation in der Seidenraupe Bombyx top (nach Leines): A – männlich, B – weiblich, C – Raupe, D – Kokon, E – Kokonpuppe
Insektenlarven mit vollständiger Metamorphose ähneln nicht den erwachsenen Larven und unterscheiden sich häufig ökologisch. So leben beispielsweise die Larven des Maikäfers im Boden und die erwachsenen Tiere auf Bäumen. Die Larven vieler Fliegen entwickeln sich im Boden, verrottendem Substrat, und erwachsene Fliegen fliegen und besuchen Blumen und ernähren sich von Nektar. Die Larven solcher Insekten häuten sich mehrmals und verwandeln sich dann in eine Puppe. In der Puppenphase kommt es zur Histolyse – der Zerstörung der Larvenorgane und zur Histogenese – der Bildung der Organisation eines erwachsenen Insekts. Aus der Puppe schlüpft ein geflügeltes Insekt – eine Imago.
So werden bei Insekten die folgenden Arten der postembryonalen Entwicklung beobachtet: Ametabolismus oder Protomorphose (Ei – Larve (ähnlich wie Imago) – Imago); Hemimetabolie – unvollständige Transformation (Ei – Nymphe – Imago): Hemimetamorphose – eine typische Variante, Hypomorphose – reduzierte Metamorphose, Hypermorphose – verstärkte Metamorphose; Holometabolie – vollständige Transformation (Ei – Larve – Puppe – Erwachsener): Holometamorphose – eine typische Variante, Hypermetamorphose – mit mehreren Arten von Larven.
Arten von Insektenlarven mit vollständiger Metamorphose. Larven holometabolischer Insekten haben im Vergleich zu erwachsenen Insekten eine einfachere Struktur. Sie haben keine Facettenaugen und keine Flügelrudimente; Der Mundapparat ist nagend, die Fühler und Beine sind kurz. Je nach Entwicklung der Gliedmaßen werden vier Larvenarten unterschieden: Protopoden, Oligopoden, Polypoden und Apoden(Abb. 342). Protopodenlarven sind charakteristisch für Bienen und Wespen. Sie haben nur Ansätze von Brustbeinen. Diese Larven sind inaktiv und entwickeln sich in Waben mit der Sorgfalt der Arbeiter. Oligopodenlarven sind häufiger als andere; sie zeichnen sich durch die normale Entwicklung von drei Laufbeinpaaren aus. Zu den Oligopoden zählen Larven von Käfern und Florfliegen. Polypodenlarven oder Raupen haben neben drei Brustbeinpaaren noch mehrere weitere falsche Beinpaare am Hinterleib. Die ventralen Crura stellen Vorsprünge des Abdomens dar
Körperwände und tragen Haken und Stacheln an der Sohle. Raupen sind charakteristisch für Schmetterlinge und Sägeblätter. Apodische oder beinlose Larven werden in der Ordnung der Diptera sowie bei einigen Käfern (Larven von Barben, Bohrern) und Schmetterlingen beobachtet.

Reis. 342. Insektenlarven mit vollständiger Metamorphose (von Barnes): A – Protopod, B, C – Oligopod, D – Polypod, E, F, G – Apod
Nach den Bewegungsmethoden werden Insektenlarven mit vollständiger Metamorphose unterteilt Campodeoid mit einem langen, flexiblen Körper, laufenden Beinen und dem Gefühl von Cerci, Erukoid mit fleischigem, leicht gebogenem Körper mit oder ohne Gliedmaßen, Draht- mit starrem Körper, rundem Durchmesser, mit tragenden Cerci - Urogomphen und wurmartig- beinlos.
Campodeoid-Larven sind charakteristisch für viele Raubkäfer – Laufkäfer, Laufkäfer. Sie bewegen sich durch Löcher im Boden. Eine typische erukoide Larve ist die Larve des Maikäfers, Mistkäfers, Bronzekäfers. Das sind grabende Larven. Drahtwürmer sind charakteristisch für Schnellkäfer und Dunkelkäfer, deren Larven aktiv durch den Boden wandern. Zahlreiche wurmartige Larven. Sie bewegen sich im Boden und im Pflanzengewebe. Dazu gehören nicht nur Diptera-Larven, sondern auch einige Käfer, Schmetterlinge und Sägeblätter, die sich beispielsweise in Pflanzengeweben entwickeln.
Arten von Puppen. Puppen sind frei, bedeckt und versteckt (Abb. 343). Bei freien Puppen sind die Rudimente von Flügeln und Gliedmaßen deutlich sichtbar und frei vom Körper getrennt, beispielsweise bei Käfern. Bei bedeckten Puppen haften alle Rudimente fest am Körper, beispielsweise bei Schmetterlingen. Die Hüllen der freien Puppen sind dünn und weich, während die der bedeckten Puppen stark sklerotisiert sind.

Reis. 343. Arten von Puppen bei Insekten (von Weber): A – freier Käfer, B – bedeckter Schmetterling, C – versteckte Fliege; 1 - Antenne, 2 - Flügelrudimente, 3 - Bein, 4 - Spirakel
Larvenhaut, die einen falschen Kokon bildet - Puparia. Im Inneren des Pupariums befindet sich eine geöffnete Puppe. Daher ist die versteckte Puppe nur eine Variante der freien. Puppen sind charakteristisch für viele Fliegen.
Oft webt die Larve im letzten Stadium vor der Verpuppung einen Kokon. So sondert beispielsweise eine Raupe eines Seidenraupenschmetterlings Seide aus den Seidendrüsen ab, aus der sie einen dichten Kokon spinnt. Im Inneren eines solchen Kokons befindet sich eine bedeckte Puppe. Und bei einigen Hymenopteren – Ameisen, sowie bei den Neuroptera – befindet sich im Kokon eine offene oder freie Puppe. Bei Florfliegenlarven, wie z. B. Goldaugenlarven, werden Fäden für den Kokon von malpighischen Gefäßen produziert und aus dem Anus ausgeschieden.
Physiologie der Metamorphose. Im Prozess der Metamorphose finden zwei miteinander verbundene Prozesse statt: Histolyse und Histogenese. Bei der Histolyse handelt es sich um den Abbau des Gewebes der Larvenorgane, bei der Histogenese um die Bildung von Organen eines erwachsenen Insekts. Bei Insekten mit unvollständiger Metamorphose laufen diese Prozesse in der Nymphenphase allmählich ab, bei Insekten mit vollständiger Metamorphose in der Puppenphase.
Die Histolyse erfolgt aufgrund der Aktivität von Phagozyten und Enzymen. In diesem Fall werden zunächst der Fettkörper, die Larvenmuskulatur und einige andere Organe zerstört, die in ein Nährsubstrat umgewandelt werden, das von sich entwickelnden Geweben verbraucht wird.
Die Histogenese oder die Bildung der Organe eines erwachsenen Insekts erfolgt hauptsächlich aufgrund der Entwicklung von Imaginalscheiben – Rudimenten aus undifferenzierten Zellen. Die Imaginalscheiben werden während der Larvenphase und sogar während der Embryogenese abgelegt und stellen innere Rudimente dar. Imaginäre Bandscheiben entwickeln Augen, Flügel, Mundapparat, Beine sowie innere Organe: Muskeln, Gonaden. Das Verdauungssystem, die malpighischen Gefäße und die Luftröhre werden nicht zerstört, sondern im Verlauf der Metamorphose stark differenziert. Das am wenigsten veränderte Herz und Nervensystem. Bei der Metamorphose im Nervensystem wird jedoch häufig der Prozess der Oligomerisierung (Fusion) von Ganglien beobachtet.
Der Prozess der Metamorphose wird durch die endokrinen Drüsen gesteuert (Abb. 329). neurosekretorische Zellen Das Gehirn schüttet Hormone aus, die die Aktivität anregen Herzkörper, deren Hormone durch die Hämolymphe stimuliert werden prothorakal(prothorakale) Drüsen, die Häutungshormone absondern - Ecdyson. Ecdyson fördert den Häutungsprozess: teilweise Auflösung und Ablösung der alten Nagelhaut sowie die Bildung einer neuen.
Im Prozess der Metamorphose ist die Aktivität benachbarte Körper produzieren jugendlich Hormon. Bei hohen Konzentrationen führt die Häutung der Larve zur Bildung einer Larve
nächstes Alter. Wenn die Larven wachsen, lässt die Aktivität der angrenzenden Körper nach, die Konzentration des Juvenilhormons sinkt und die Brustdrüsen degenerieren allmählich. Dies führt dazu, dass sich die Larven in die Puppenphase und dann in die adulte Phase häuten.
Die künstliche Transplantation benachbarter Körper, beispielsweise in eine Heuschreckennymphe des letzten Alters, trägt dazu bei, dass sie sich nicht in die Erwachsenenphase, sondern in eine größere Larve zusätzlichen Alters häutet. In der Erwachsenenphase steuert das Juvenilhormon die Entwicklung der Gonaden und das Hormon Ecdyson wird aufgrund der Verkleinerung der Prothorakaldrüsen nicht mehr produziert.
Ursprung der Metamorphose. Über den Ursprung der Metamorphose bei Insekten gibt es mehrere Hypothesen. Lange Zeit gab es Streit darüber, welche Insekten evolutionär weiter fortgeschritten sind – mit vollständiger oder unvollständiger Transformation. Einerseits sind Insektennymphen mit unvollständiger Metamorphose fortgeschrittener entwickelt als Insektenlarven mit vollständiger Metamorphose, andererseits haben letztere eine fortgeschrittene Puppenphase.
Gegenwärtig wurde dieser Widerspruch durch die Hypothese des Ursprungs der Metamorphose von G. S. Gilyarov, A. A. Zakhvatkin und A. G. Sharov beseitigt. Nach dieser Hypothese entwickelten sich beide Formen der Metamorphose bei Insekten unabhängig von einer einfacheren Art der Entwicklung – der Protomorphose, die bei primären flügellosen Insekten, beispielsweise bei Insekten mit Borstenschwanz (Thysanura), beobachtet wird.
Bei der Protomorphose erfolgt die Entwicklung direkt, wobei viele Häutungen in der Larvenphase und dann im Imaginalstadium beobachtet werden. Alle Entwicklungsphasen dieser Insekten finden in derselben Umgebung statt.
Es wird angenommen, dass Insekten im Laufe der Evolution von einer halbversteckten Existenz in der oberen Bodenschicht zu einem Leben auf der Oberfläche und auf Pflanzen übergegangen sind. Dieser Übergang in einen neuen Lebensraum wurde von einer großen Aromorphose gekrönt – der Entwicklung von Flügeln und Flug.
Die Entwicklung offener Lebensräume spiegelte sich in der Entwicklung der individuellen Entwicklung von Insekten wider. Die Evolution der Insektenontogenese verlief offenbar zwei Hauptrichtungen.
In einem Fall war der Prozess der Embryonisierung der Entwicklung im Gange, was dazu führte, dass in späteren Entwicklungsphasen Insekten aus dotterreichen Eiern schlüpften. Dies führte zur Vorstellung von Larven mit der Bildung von Nymphen. So entstanden Insekten mit unvollständiger Metamorphose. Dieser Evolutionsweg führte zur fortschreitenden Entwicklung von Larven, die eine ähnliche Lebensweise wie Erwachsene führten.
Im anderen Fall fand dagegen ein Prozess der Deembryonisierung der Entwicklung statt, d. h. es wurden dotterarme Eier in früheren Entwicklungsstadien ausgebrütet. Dies führte zur morphoökologischen Divergenz von Insektenlarven und adulten Insekten. Die Larven haben sich vereinfacht und angepasst
in einer geschützteren Umgebung zu leben und die Funktion der Ernährung zu erfüllen, und Erwachsene begannen, hauptsächlich die Funktion der Fortpflanzung und Ansiedlung zu erfüllen. Neben der Deembryonisierung der Entwicklung von Insektenlarven mit vollständiger Transformation haben sie viele vorläufige Anpassungen an verschiedene Existenzbedingungen entwickelt. So kam es in der Evolution von Larven und Erwachsenen bei Insekten mit vollständiger Metamorphose zu Amphigenese (Divergenz). Die Amphigenese von Larven und Erwachsenen erwies sich hinsichtlich der morphologischen Anpassungen als sehr tiefgreifend, was zu ernsthaften Widersprüchen in der Ontogenese führte. Sie wurden durch das Aufkommen der Puppenphase erfolgreich gelöst, in der eine radikale Umstrukturierung der Larvenorganisation hin zur Imaginalphase stattfindet. Dadurch konnten Insekten mit vollständiger Transformation ein breiteres Spektrum ökologischer Nischen erobern und ein beispielloses Aufblühen unter den Tieren auf der Erde erreichen.
Insektenzucht. Für die meisten Insekten ist die bisexuelle sexuelle Fortpflanzung charakteristisch. Viele Arten weisen einen Sexualdimorphismus auf. Beispielsweise haben männliche Hirschkäfer zu Hörnern umgestaltete Mandibeln, und männliche Nashornkäfer haben ein Horn auf dem Kopf und Höcker auf dem Halsschild. Dies ist auf das Paarungsverhalten dieser Arten zurückzuführen, das mit dem Kampf der Männchen um ein Weibchen einhergeht. Die Beziehung zwischen den Geschlechtern verschiedener Arten ist äußerst vielfältig. Männliche Dipteren aus der Familie der Dolichopodidae bringen dem Weibchen ein „Geschenk“ – eine gefangene Fliege und führen einen Tanz mit Beinspiegeln auf. Gottesanbeterinweibchen sind räuberisch veranlagt und fressen das Männchen während der Paarung.
Die meisten Insekten legen Eier, es werden jedoch häufig Lebendgeburten beobachtet. In diesem Fall entwickeln sich die Eier im weiblichen Genitaltrakt und sie bringt Larven zur Welt. Schmeißfliegen-Sarkophage (Sarcophagidae) legen beispielsweise lebende Larven auf das Fleisch, deren Entwicklung sehr schnell voranschreitet. Kein Wunder, dass man in der Antike glaubte, dass Würmer im Fleisch spontan entstehen. Man kann nicht bemerken, wie eine Schmeißfliege offen liegendes Fleisch besucht, und plötzlich weiße Larven entdecken, die plötzlich auftauchten.
Zu den lebendgebärenden Arten zählen auch die Schafsblutsaugerfliege, einige Käfer, die in Höhlen leben.
Zusätzlich zur bisexuellen sexuellen Fortpflanzung weisen einige Insekten eine Parthenogenese auf – eine Entwicklung ohne Befruchtung. Es gibt viele Arten aus verschiedenen Insektenordnungen, die durch Parthenogenese gekennzeichnet sind. Die Parthenogenese kann obligat sein – obligatorisch, dann sind alle Individuen der Art nur Weibchen. So kommen unter alpinen Bedingungen, im Norden und unter anderen ungünstigen Bedingungen parthenogenetische Käfer, Orthopteren, Ohrwürmer und Florfliegen vor. Parthenogenese findet auch bei bisexuellen Arten statt, wenn ein Teil der Eier befruchtet und ein Teil ohne Befruchtung gelegt wird. Bienendrohnen entwickeln sich beispielsweise aus unbefruchteten Eiern.
Eine ähnliche Parthenogenese findet sich auch bei anderen Hymenopteren (Ameisen, Sägeblätter), Termiten, einigen Wanzen und Käfern. Und bei Blattläusen gibt es beispielsweise einen Generationswechsel im Lebenszyklus: bisexuell und parthenogenetisch. In einigen Fällen kann die Parthenogenese optional (vorübergehend) sein und sich nur unter widrigen Bedingungen manifestieren. Die Parthenogenese bei Insekten trägt zur Erhaltung hoher Populationen bei.
Eine Variante der Parthenogenese ist die Pedogenese – Fortpflanzung ohne Befruchtung in der Larvenentwicklungsphase. Dies ist eine besondere Art der Entwicklung von Insekten, bei der die Reifung der Keimdrüsen der anderer Organe voraus ist. Einige Arten von Gallmücken vermehren sich beispielsweise in der Larvenphase. Ältere Larven bringen jüngere Larven zur Welt. Bei einer der Käferarten wurde Pedogenese festgestellt, deren Larven teilweise Eier legen und teilweise Larven zur Welt bringen. Die Pedogenese erhöht zusätzlich zur bisexuellen Fortpflanzung die Artenvielfalt.
Lebenszyklen von Insekten. Im Gegensatz zur Ontogenese bzw. der individuellen Entwicklung von Insekten handelt es sich beim Lebenszyklus um die Entwicklung einer Art, die in der Regel mehrere Arten von Ontogeneien umfasst. Die Ontogenese beschränkt sich auf das Leben eines Individuums von der Eizelle über den Beginn der Pubertät bis hin zum natürlichen Tod. Der Lebenszyklus ist ein wiederkehrender Teil der kontinuierlichen Entwicklung einer Art. So besteht der Lebenszyklus im typischsten Fall bei Insekten aus zwei konjugierten und morphophysiologisch unterschiedlichen Ontogenesen von Männchen und Weibchen, die sich sexuell vermehren und ihresgleichen vermehren. Und bei parthenogenetischen Arten ist der Lebenszyklus nur durch eine Art weiblicher Ontogenese gekennzeichnet.
Die Lebenszyklen von Insekten sind hinsichtlich der Art der Fortpflanzung, der Zusammensetzung der Generationen und ihres Wechsels vielfältig. Die folgenden Arten von Insektenlebenszyklen können unterschieden werden.
1. Lebenszyklen ohne Generationswechsel mit bisexueller sexueller Fortpflanzung. Dies ist der häufigste Lebenszyklustyp, der für dimorphe Arten charakteristisch ist und nur aus Männchen und sich sexuell fortpflanzenden Weibchen besteht. Dies sind die Zyklen der meisten Käfer, Schmetterlinge und Wanzen.
2. Lebenszyklen ohne Generationswechsel mit parthenogenetischer Fortpflanzung. Solche Arten sind monomorph und bestehen nur aus parthenogenetischen Weibchen, die ohne Befruchtung Eier legen.
Parthenogenetische Arten kommen besonders häufig bei Blattläusen, Blattläusen und anderen Homopteren vor. Parthenogenetische Arten von Käfern, Wanzen, Heuschrecken und Kokziden kommen im Hochgebirge häufig vor.
3. Der seltenste Lebenszyklustyp bei Insekten ist ein Zyklus ohne Generationswechsel mit sexueller Fortpflanzung zwittriger Arten.
Bekannt ist eine amerikanische Fliegenart, die nur aus zwittrigen Individuen besteht. In den frühen Phasen der Entwicklung fungieren die erwachsenen Tiere als Männchen, in späteren Stadien als Weibchen. Daher legen alle Individuen Eier, was die Artenvielfalt erhöht.
4. Lebenszyklen ohne Generationswechsel mit sexueller Fortpflanzung und fakultativer Parthenogenese bei polymorphen Arten, beispielsweise bei sozialen Insekten. Die Art besteht aus sexuellen Individuen – Männchen und Weibchen sowie fruchtbaren – arbeitenden Individuen, die nicht an der Fortpflanzung teilnehmen. Zu diesen Arten gehören Bienen, Ameisen und Termiten. Solche Lebenszyklen werden durch die Tatsache erschwert, dass Weibchen neben befruchteten Eiern auch parthenogenetische Eier legen, aus denen beispielsweise Bienen haploide Männchen – Drohnen – und aus befruchteten Eiern Weibchen und Arbeiterinnen entwickeln. Einige Reiter, Thripse und Kokziden entwickeln sich ähnlich.
Bei anderen Arten manifestiert sich die fakultative Parthenogenese auf andere Weise: Aus unbefruchteten Eiern entwickeln sich nicht Männchen, sondern Weibchen. Aber in diesem Fall wird der diploide Chromosomensatz bei Frauen durch die Fusion haploider Kerne wiederhergestellt. Eine solche Entwicklung ist bei einigen Stabheuschrecken, Heuschrecken, Blattwespen und Kokziden bekannt.
5. Lebenszyklen mit Wechsel der Geschlechtsgeneration und Parthenogenetik (Heterogonie). Bei vielen Blattläusen und Reblausen gibt es neben der sexuellen Generation geflügelter Männchen und Weibchen mehrere abwechselnde Generationen parthenogenetischer Weibchen, geflügelt oder flügellos.
6. Lebenszyklen mit Wechsel der Geschlechtsgeneration und mehreren Generationen mit Pedogenese. Beispielsweise kommt es bei einigen Gallmücken nach der sexuellen Fortpflanzung, an der Männchen und Weibchen beteiligt sind, zur parthenogenetischen Fortpflanzung der Larven (Pedogenese). Nach mehreren Generationen brütender Larven, die nach der Lebendgeburt ihrer Artgenossen absterben, verpuppt sich die letzte Larvengeneration und bringt geflügelte Weibchen und Männchen hervor.
7. Lebenszyklen mit Wechsel der sexuellen Generation (männlich und weiblich) mit asexueller. Nach der sexuellen Fortpflanzung legen die Weibchen befruchtete Eier, die eine Polyembryonie durchlaufen. Dies ist eine ungeschlechtliche Fortpflanzung in der Embryonalphase. Das Ei wird zerdrückt und der Embryo in der Morula-Phase beginnt sich durch Knospung zu vermehren. Aus einer Eizelle können mehrere Dutzend Embryonen entstehen. Solch
Somit lässt sich die Einteilung der Lebenszyklen wie folgt darstellen.
ICH. Kein Generationswechsel:
- 1) mit bisexueller sexueller Fortpflanzung (Maikäfer);
- 2) mit parthenogenetischer Fortpflanzung (Alpenkäfer, Heuschrecken);
- 3) mit sexueller Fortpflanzung hermaphroditischer Individuen (Amerikanische Fliege);
- 4) mit sexueller Fortpflanzung und teilweiser Parthenogenese bei polymorphen Arten (Bienen).
II. Mit Generationswechsel:
- 1) Heterogonie: Wechsel der Geschlechtsgeneration und mehrerer parthenogenetischer (Blattläuse, Reblaus);
- 2) Heterogonie: Wechsel der sexuellen Generation und mehrerer pädogenetischer Generationen (einige Gallmücken);
- 3) Metagenese: Wechsel der sexuellen Generation mit Polyembryonie (Reiter).
Saisonale Zyklen von Insekten. Versteht man den Lebenszyklus als einen sich zyklisch wiederholenden Teil der Morphogenese einer Art von einer Entwicklungsphase zur gleichen Phase, so versteht man unter dem saisonalen Entwicklungszyklus ein Merkmal der Entwicklung einer Art während der Jahreszeiten eines Jahres ( von Winter zu Winter).
So dauert beispielsweise der Lebenszyklus des Maikäfers 4-5 Jahre (vom Ei bis zum reifen Erwachsenen), und der saisonale Zyklus dieser Art ist dadurch gekennzeichnet, dass sich überwinternde Larven im Frühjahr verpuppen und junge Käfer sich vermehren. Im Sommer, Herbst und Winter sind ihre Larven unterschiedlichen Alters zu finden. Die Anzahl der Generationen, die sich im Laufe eines Jahres entwickeln, wird Voltness genannt.
Es gibt Arten, die mehrere Generationen pro Jahr hervorbringen. Dies sind multivoltaische Arten. Beispielsweise kann eine Stubenfliege pro Saison 2-3 Generationen hervorbringen und überwintert im Erwachsenenalter. Die meisten Insekten sind monovoltin und produzieren eine Generation pro Jahr.
Saisonale Zyklen von Insekten in der Natur sind durch kalendarische Zeiträume des Auftretens verschiedener Entwicklungsphasen gekennzeichnet. Wichtige Merkmale der saisonalen Zyklen von Arten sind der Zeitpunkt ihres aktiven Lebens und die Diapause (vorübergehende Verzögerung der Entwicklung) im Winter oder Sommer. Die Regulierung der Lebenszyklen von Arten entsprechend lokalen saisonalen Phänomenen erfolgt durch Umweltfaktoren und das neurohumorale System des Körpers.
Die embryonale bzw. embryonale Entwicklung der Arbeitsbiene, der Gebärmutter, der Drohne umfasst alle Veränderungen, die unter der Eierschale stattfinden, wodurch aus einem einzelligen Ei ein vielzelliges Lebewesen, die Larve, entsteht (Abb. 32). ). Das gerade von der Gebärmutter gelegte Ei wird in der Regel mit einem Ende am Zellboden festgeklebt und steht senkrecht darauf. Die Länge des Eies beträgt 1,6–1,8 mm, die Breite 0,31–0,33 mm. Es ist leicht gebogen, von länglicher zylindrischer Form, das der Befestigungsstelle gegenüberliegende Ende ist leicht erweitert. An
- Am 3. Tag nimmt das Ei eine Schräglage ein und liegt am 3. Tag auf dem Zellboden. Am freien, ausgebreiteten Ende der Eizelle befindet sich ein kleines Loch, durch das beim Durchgang der Eizelle durch den Eileiter Spermien aus dem Uterusgefäß eindringen. Dieses Loch wird Mikropyle genannt. Zu anderen Zeiten ist die Mikropyle sowohl für Bakterien als auch für Viren undurchlässig. Das Ei hat ein vorderes und ein hinteres Ende. Der Kopf der Larve entwickelt sich im freien vorderen Ende des Eies.
Die Embryonalentwicklung beginnt mit der Fragmentierung des Zellkerns. Die durch die Teilung gebildeten Tochterkerne befinden sich zunächst zufällig im Eigelb, wandern dann mit kleinen Bereichen des Zytoplasmas an die Peripherie des Eies und bilden eine durchgehende Zellschicht – das Blastoderm, das die gesamte Eischale auskleidet das Innere. Auf der konvexen Seite des Eies beginnen Blastodermzellen schneller zu wachsen und sich zu vermehren als die Zellen auf der konkaven Seite. Dadurch entsteht ein Embryonalstreifen in Form eines breiten Strangs. Dies ist der Beginn der Bildung des Embryos. Der Keimstreifen wächst aufgrund der Zellteilung und -differenzierung sowie der Gewebebildung. Als Ergebnis dieser Prozesse kommt es zur Entwicklung der Larve. In der zweiten Hälfte des 2. Tages beginnt der Prozess der Segmentierung des Embryos. Am vorderen Ende des Keimstreifens bildet sich ein Präorallappen (Akron), auf dem sich ein Tuberkel bildet – das Rudiment der Oberlippe. Unterhalb dieses Tuberkels erscheint eine Aussparung – der zukünftige Mund. Dann - die Rudimente der Antennen. Die Segmente des Brustkorbs und des Abdomens lösen sich nach hinten vom Kopfsegment. Von der ventralen (ventralen) Seite dieser Segmente erscheinen Auswüchse – die Rudimente gepaarter Gliedmaßen. Zuerst tritt es auf der Brust auf, später am Bauch. Gleichzeitig findet sich im Kopfbereich eine deutliche Segmentierung: Es bilden sich die Rudimente von fünf Kopfsegmenten aus. Darüber hinaus erscheinen Atemlöcher – Narben, sowie Anfänge der Spinndrüsen und Malpighischen Gefäße. Bei der Bildung der Körper- und Gliedmaßensegmente kommt es zur Trennung einzelner Teile.
Anschließend vergrößert sich der Keimstreifen. Die Vermehrung von Blastodermzellen führt zur Einstülpung des mittleren Teils des Keimstreifens (Gastrulation), der tiefer in die Eizelle eindringt und sich vom Blastoderm trennt. Bei der Gastrulation bildet sich zunächst eine Längsrille auf der Oberfläche des Keimstreifens, und dann beginnen die Außenkanten der Rille aufeinander zu zu wachsen. Dadurch stülpt sich die zukünftige zweite Keimschicht in den Keimstreifen ein. Wenn sich die Ränder der Furche schließen, entsteht die äußere Schicht (Ektoderm). Darunter liegt die innere Schicht (Mesoderm). An den Enden der Mesodermschicht treten Einstülpungen auf, die aus Zellgruppen bestehen, die die dritte Keimschicht (Endoderm) bilden. Die Rudimente einzelner Organe beginnen zu erscheinen. Ihre Entwicklung und Differenzierung setzt sich fort, bis die Larve aus der Zelle schlüpft.
Aus dem Ektoderm werden die Körperwand, der vordere und hintere Darm, das Luftröhren- und Nervensystem sowie viele Drüsen gebildet; aus dem Endoderm - die Rudimente des Stachels, der Flügel und Gliedmaßen, die Endabschnitte des Genitaltrakts, des Nervensystems und der Sinnesorgane; aus dem Mesoderm - Muskeln, Fettkörper, Geschlechtsdrüsen.
Ein vollständig ausgebildeter Embryo, der sich im Wesentlichen in eine Larve verwandelt hat, füllt das gesamte Ei aus. Sie beginnt sich intensiv zu bewegen, Luft in die Luftröhre zu saugen, Fruchtwasser zu schlucken und dadurch ihr Körpervolumen zu vergrößern. Die Larve wird aus den Embryonalmembranen freigesetzt, durchbricht das Chorion (die Sekundärhülle des Eies, die durch die Transformation der Follikelepithelzellen entsteht) und schlüpft, das heißt, sie kommt heraus. Normalerweise dauert die Embryonalentwicklung von Honigbienen etwa 3 Tage. Temperaturänderungen können die Dauer der Embryonalentwicklung verlängern oder verkürzen.
Nach der Freisetzung der Larve aus dem Ei beginnt die postembryonale Entwicklung, die mit einer Metamorphose (Transformation) einhergeht.
Metamorphose ist eine tiefgreifende Veränderung der Struktur des Organismus, durch die sich die Larve in ein erwachsenes Insekt verwandelt. Abhängig von der Art der postembryonalen Entwicklung bei Insekten werden zwei Arten der Metamorphose unterschieden:
unvollständig (Hemimetabolismus), wenn die Entwicklung eines Insekts durch den Durchgang von nur drei Stadien gekennzeichnet ist – Eier, Larven und die Erwachsenenphase (Erwachsene);
vollständig (Holometabolismus), wenn der Übergang der Larve in die erwachsene Form in einem Zwischenstadium erfolgt - dem Puppenstadium.
Bei unvollständiger Transformation ist die Lebensweise der frühen Stadien und der Erwachsenen ähnlich. Die Larve mit dieser Art der Entwicklung ähnelt einem erwachsenen Insekt und verfügt wie dieses über Facettenaugen, Mundorgane und dann von außen deutlich sichtbare Flügelrudimente. Die Flügelrudimente nehmen mit jeder weiteren Häutung an Größe zu und können im letzten präimaginären Alter bereits mehrere Abschnitte des Hinterleibs bedecken. Bei einem Insekt im letzten präimaginären Alter sind die Imaginalstrukturen vollständig entwickelt, und als Ergebnis der letzten Häutung erscheint ein vollständig ausgebildeter Erwachsener. Eine unvollständige Transformation ist charakteristisch für Orthopteren, Käfer, Homopteren usw.
Bei vollständiger Metamorphose ist der gesamte Entwicklungszyklus durch den Übergang der Stadien Ei, Larve, Präpuppe, Puppe und Erwachsener gekennzeichnet (Tabelle 3).
- Die Dauer der Entwicklungsstadien von Individuen des Bienenvolkes, Tage
Der Übergang von der Larve zum erwachsenen Tier erfolgt im Puppenstadium. Dies ist ein nicht nährendes Stadium, in dem Gewebe dazugehören
In früheren Stadien bilden sich imaginäre Strukturen aus. Bei den meisten Insekten ist dieses Stadium unbeweglich. Die Puppe reagiert nicht auf äußere Reize und ist mit einer dicken Decke bekleidet. Am Ende dieser Phase platzt die Schale und ein erwachsenes Insekt erscheint.
Bei Insekten erfolgt die Regulierung der Metamorphose durch Hormone, die von den endokrinen Drüsen (endokrinen Drüsen) produziert werden. Derzeit sind drei Metamorphosehormone bekannt, die die postembryonale Entwicklung steuern: Juvenilhormon, Häutungshormon (Larve) und Aktivierungshormon. Juvenile Hormone werden in den angrenzenden Körpern produziert und gespeichert. Hierbei handelt es sich um zwei kleine Zellstrukturen, die hinter dem Gehirn an den Seiten der Speiseröhre liegen.
Wenn einer jungen Larve die angrenzenden Körper entfernt werden, erfolgt nach der nächsten Häutung die Verpuppung, obwohl sie im Normalzustand noch mehrere Häutungen durchführen müsste. Dieses Hormon bestimmt das Wachstum der Larven und steuert die Entwicklung einer Reihe von Organen und Prozessen. Beispielsweise kann der Fettkörper ohne dieses Hormon nicht wachsen und nicht funktionieren.
Das Häutungshormon oder Ecdyson wird in den Prothorakaldrüsen produziert, die sich im Prothorax der Larve oberhalb des ersten Nervenganglions in der Nähe des ersten Spirakels befinden. Es wird im Körper des Insekts synthetisiert. Cholesterin. Das Hormon löst den Häutungsprozess aus und reguliert so indirekt das Wachstum und die Strukturbildung. Die Einführung dieses Hormons bei Insekten führt zur Bildung von Schwellungen auf Riesenchromosomen. Die Analyse dieser Schwellungen ergab, dass in ihnen ein intensiver Prozess der RNA-Bildung stattfindet. Es wird angenommen, dass das erste Ergebnis der Wirkung von Ecdyson die Aktivierung von Genen, dann die Stimulierung der RNA-Biosynthese und die Bildung der entsprechenden Enzyme ist, die den Prozess der Metamorphose sicherstellen.
Das aktivierende Hormon wird in speziellen neurosekretorischen Zellen produziert, die sich im vorderen dorsalen Teil des supraösophagealen Ganglions befinden. Diese Drüsen sind bei Arbeiterbienen besser entwickelt, bei Königinnen weniger und bei Drohnen noch weniger. Das Aktivierungshormon beeinflusst die Wiederaufnahme der Insektenaktivität nach jeder Häutung und stimuliert die Bildung von zwei weiteren Hormonen – Ecdyson und Juvenil.
Die Honigbiene ist ein Insekt mit vollständiger Metamorphose (Abb. 33).
Larvenstadium. Es zeichnet sich durch intensive Ernährung und Wachstum aus. Das Stadium der Arbeiterbienenlarve dauert 6 Tage, das der Königin 5 und das der Drohne 7. In den ersten Tagen erhalten die Larven der Honigbienen so viel Nahrung, dass sie auf der Oberfläche schwimmen. Die Grundlage des Futters ist das Geheimnis von Hypofarin-
| 1 g | 3 | 4 5 | 6 7 8 Ampere; | 10 | 11 12 | 13 14 15 16 17 18 19 20 | 21 |
| Sch |
Tag
Reis. 33. Stadien der Bienenentwicklung
Zahndrüsen von Ammenbienen – eine durchscheinende weiße Flüssigkeit. Die von den Drüsen der Ammenbienen abgesonderte Milch ist sehr nahrhaft. Es enthält Kohlenhydrate, Proteine, Fette, Mineralien und Vitamine der Gruppe B. Bevor die Arbeitsbiene der Larve Nahrung gibt, senkt sie ihren Kopf in die Zelle und öffnet und schließt wiederholt den Oberkiefer. Dann berührt es mit seinen Kiefern den Boden der Zelle und gibt flüssige Nahrung ab. Andere Ammenbienen geben auf die gleiche Weise die folgenden Portionen Milch in die Zelle mit der Larve. Während des gesamten Larvenstadiums gibt es etwa 10.000 Besuche einer Zelle mit einer Larve.
Die Nahrung junger Larven und älterer Larven ist nicht gleich. Im zweiten Lebensabschnitt beginnt die Larve der Arbeitsbiene mit der Aufnahme von Brei – einer Mischung, die von Ammenbienen aus Honig und Pollen zubereitet wird.
Während des Larvenstadiums vergrößern sich die linearen Abmessungen der Arbeiterbienenlarven um mehr als das Zehnfache. So beträgt die Länge einer frisch geschlüpften Larve 1,6 mm, einer 1 Tag alten Larve 2,6 mm, einer 2 Tage alten Larve 6 mm und am Ende des Larvenstadiums 17 mm (sie nimmt sie fast vollständig ein). am Boden der Zelle). Die Länge der Uteruslarve erreicht am Ende des Stadiums 26,5 mm.
Durch eine intensive Ernährung kommt es bei den Larven zu einer deutlichen Gewichtszunahme. Die geschlüpfte Larve wiegt etwa 0,1 mg. Zum ersten Mal
- Tage erhöht sich die Masse einer Arbeiterbienenlarve um das 45-fache und die Masse einer Drohnenlarve um das 85-fache. Am Ende des Stadiums erhöht sich das Gewicht der Larven arbeitender Individuen um das 1565-fache und das der Mütter um das 2926-fache.
Im Aussehen unterscheidet sich die Larve stark von der erwachsenen Biene (Abb. 34). Es hat eine wurmartige Form und weiche, weißliche Hüllen. Sein Körper besteht aus einem Kopf, einem gegliederten Rumpf und einem Anallappen.
Der Kopf der Larve ist klein, stumpfkegelförmig, die Basis des Kegels wird von der Kopfkapsel gebildet, die durch eine Längsnaht geteilt ist.
Reis. 34. Äußere (G) und innere (1G) Struktur der Bienenlarve "\u003e
Reis. 34. Äußere (D) und innere (1D) Struktur der Bienenlarve:
A – Gesamtansicht; B – Ansicht von der Seite des Kopfes; C – Position der erwachsenen Larve am Boden der Zelle; G – Seitenansicht; y - Antennenrudimente; vg – Oberlippe; vh – Oberkiefer; opzh - Öffnung der Spinndrüse; ng – Unterlippe; p - Mund; ndg – supraösophageales Ganglion; PC – Vorderdarm; a - Aorta; sk - Mitteldarm; zht – fetter Körper; zpzh – der Keim der Geschlechtsdrüse; c – Herz; zk – Hinterdarm; ms – malpighisches Gefäß; bn – Bauchnervenkette; pzh - Eisenspinnerei; pdg – subösophageales Ganglion
in zwei gleiche konvexe Teile, die sogenannten Wangen. Die Spitze des Kegels an der Vorderseite des Kopfes wird von der Oberlippe und anderen Mundwerkzeugen gebildet. Der Teil des Kopfes, einschließlich der Unterlippe, des Oberkiefers und des Teils der Kopfkapsel, an dem sie befestigt sind, ist von der Kopfkapsel durch eine tiefe Rille getrennt, die auf beiden Seiten zwischen der Basis des Oberkiefers verläuft und Oberkiefer und erstreckt sich bis zu den hinteren Rändern des Kopfes.
Die Mandibeln erstrecken sich in Form kegelförmiger Organe von der Kopfkapsel. Die Antennenrudimente befinden sich in Form runder Tuberkel auf der Kopfkapsel über der Basis der Mandibeln.
Die Larven an den Brustsegmenten haben keine äußeren Flügel- oder Beinansätze. Es gibt keine Facettenaugen. Unter der Larvenkutikula sind jedoch ihre Rudimente angedeutet, die als Imaginalknospen bezeichnet werden. Die Imaginalknospen der Beine sind spindelförmig und werden durch Querrillen präpariert. Die Rudimente der Flügel bestehen aus zwei flachen, nach innen gebogenen Fortsätzen der Unterhaut. Sie befinden sich an den Seiten des zweiten und dritten Brustsegments. In der Region Vom achten und neunten Abdomensegment kommen auf der Bauchseite die Rudimente der Geschlechtsorgane zum Vorschein.
Es ist zu beachten, dass die Entwicklung von Flügeln, Fühlern, Augen und Beinen nicht durch die Umstrukturierung reduzierter Larvenstrukturen erfolgt; sie entwickeln sich aus isolierten Bereichen embryonalen Gewebes, die sich in einem inaktiven Zustand befinden und als Rudimente bezeichnet werden.
Die Larven unterscheiden sich von der erwachsenen Biene durch den Aufbau der inneren Organe (siehe Abb. 34). Den zentralen Platz im Körper der Larve nimmt der Darmkanal ein, der aus drei Abschnitten besteht – dem Vorder-, Mittel- und Hinterdarm. Der Vorderdarm sieht aus wie ein kurzer, dünner Schlauch, der am hinteren Ende leicht erweitert ist. Der vordere Darm enthält eine kleine Mundhöhle, einen kurzen Rachen und eine Speiseröhre. Der für erwachsene Bienen charakteristische Honigstrupf fehlt. In der Wand des Vorderdarms befinden sich Muskeln, die für die Aufnahme von flüssigem Futter sorgen. An der Verbindung von Speiseröhre und Mitteldarm befindet sich eine kleine Ringfalte, die als Klappe fungiert. Es verschließt das Lumen der Speiseröhre und verhindert, dass der Inhalt des Mitteldarms in die Speiseröhre zurückfließt. Am Ende des Larvenstadiums bildet sich anstelle der Klappe ein dickes Septum, das den Mitteldarm bedeckt. 1
Der Mitteldarm ist der größte Abschnitt des Darms und nimmt den größten Teil des Körpers der Larve ein. Eine Besonderheit dieses Abschnitts ist das Fehlen seiner Verbindungen zum Hinterdarm, wodurch unverdaute Teile der Nahrung während des gesamten Larvenstadiums darin verbleiben. Die Verbindung dieser beiden Abschnitte erfolgt erst vor dem Spinnen des Kokons, wenn die Larve bereits versiegelt ist. Während dieser Zeit bricht die Membran, die den Mitteldarm vom Hinterdarm trennt, durch und der Kot gelangt in den Hinterdarm und wird anschließend aus dem Körper bis zum Zellboden entfernt.
Im Mitteldarm besitzen die Larven malpighische Gefäße. Die Honigbiene hat vier davon. Es handelt sich um lange, leicht gewundene Röhren, die entlang des Mitteldarms verlaufen. Während des Larvenlebens sind die malpighischen Gefäße geschlossen. Erst am 6. Tag des Larvenstadiums, wenn sie durch die dort angesammelten Zerfallsprodukte stark anschwellen, brechen die hinteren Enden und der Inhalt durch, gelangen in den Hinterdarm und werden dann entfernt. Daher entleert sich die Larve während ihres Wachstums und der Nahrungsaufnahme nicht.
Der Hinterdarm ist klein und eine schmale Röhre, die in einem spitzen Winkel gebogen ist und im Anus endet.
Das Herz der Larve besteht aus 12 Kammern gleicher Struktur (bei einer erwachsenen Biene sind es nur fünf) und ist eine dünnwandige Röhre, die sich direkt unter der Kutikula von Bauch und Brust befindet. Am zweiten Segment der Brustregion verläuft die Aorta. Es sieht aus wie eine nach unten gebogene Röhre, die dann in den Kopf übergeht und an der Vorderfläche des Gehirns endet, wo die Hämolymphe in die Körperhöhle strömt. Auf der Oberfläche der Aorta befindet sich ein dichtes Netz von Luftröhren.
Zwischen allen Herzkammern gibt es Öffnungen, durch die die Hämolymphe in das Herz gelangt. Aufgrund der Tatsache, dass die dünnen Vorderenden jeder Kammer in die nächste Kammer hineinragen, ist kein Rückfluss der Flüssigkeit möglich.
Das Atmungssystem der Larve ist recht einfach und wird durch zwei große Längstrachealstämme dargestellt, die an den Seiten des Körpers verlaufen und von denen sich kleine Äste über den ganzen Körper erstrecken. An den Seiten der Segmente befinden sich 10 Paar Stigmen, bei denen es sich um Hohlräume handelt, die von schmalen Chitinringen umgeben sind.
Das Nervensystem besteht aus zwei einfach angeordneten Kopfganglien (einem großen supraösophagealen und einem kleinen subglottalen Ganglien) und einem ventralen Nervenstrang, der sich entlang der Unterseite von Brust und Bauch erstreckt. Die Bauchkette besteht aus 11 Ganglien – 3 Brustganglien und 8 Bauchganglien. Das letzte Bauchganglion liegt in der Mitte des elften Segments; Es entstand durch die Verschmelzung der Ganglien der letzten drei Segmente. Die Ganglien sind durch paarige Konnektivitäten miteinander verbunden. Das Ganglion subösophageale ist außerdem über zwei Konnektoren mit dem ventralen Nervenstrang verbunden.
Die Geschlechtsorgane sind in der Larve der Arbeiterbiene bereits im Säuglingsalter vorhanden und werden durch zwei Leisten im achten Segment dargestellt. Ihre Länge beträgt durchschnittlich 0,27 mm und ihre Breite 0,14 mm. Während des gesamten Larvenstadiums kommt es zur Entwicklung von Ovarialprimordien. Bei der 2 Tage alten Larve sind sie von unbedeutender Größe, bei der 6 Tage alten Larve sind sie bereits verlängert und befinden sich bis zum kaudalen Ende bis zum neunten Segment. Am Ende des Larvenstadiums erreicht die Zahl der Eiröhren einer Arbeitsbiene 130–150. In den folgenden Stadien, wenn sich die Larve in eine Puppe verwandelt, zerfällt das Larvengewebe und die Anzahl der Eileiter sinkt bei der Arbeitsbiene von 20 auf 3, während sich in der Gebärmutter die Eierstöcke im Puppenstadium weiterentwickeln.
Der Fettkörper ist in der Larve gut entwickelt, wo es zur Anreicherung von Nährstoffen kommt. Bei älteren Larven erreicht der Fettkörper 60 % des Körpergewichts und füllt fast den gesamten Hohlraum zwischen Mitteldarm und Kutikula der Larve aus. Der Fettkörper ist eine Konzentration von Fetten, Proteinen und Kohlenhydraten, aufgrund derer sich in der Zukunft die Puppe entwickelt. Der Fettkörper besteht neben den eigentlichen Fettzellen auch aus Ausscheidungszellen und Enozyten, die Harnsäuresalze einfangen.
Die Spinndrüse ist nur bei den Larven vorhanden. Das Sekret dieser Drüse dient als Material zum Spinnen des Kokons vor dem Übergang der Larve in das Puppenstadium. Die Spinndrüse sieht aus wie zwei lange Röhren, die von der Bauchseite unter den Mitteldarm gelegt werden. Beim vorderen Chag sind beide Röhren zu einem unpaarigen Ausführungsgang verbunden, der unter der Mundöffnung an der Spitze der Unterlippe mündet.
Während des gesamten Stadiums entwickelt die Larve Rudimente eines Stachels, von denen sich ein Paar auf dem elften Segment und zwei Paare auf dem zwölften befinden. Zwischen ihnen befindet sich segmentweise ein Rudiment der Genitalöffnung. Das Rudiment einer großen Giftdrüse erscheint erst gegen Ende des Larvenstadiums, und zu Beginn des Puppenstadiums bildet sich aus der Vertiefung der Hülle am neunten Segment eine kleine Giftdrüse.
Am 5.-6. Tag nach der Freisetzung der Larven aus dem Ei beginnen die Bienen, die Zelle mit einer Wachskappe zu verschließen. Die Zusammensetzung des Materials, aus dem die Kappe besteht, umfasst neben Wachs auch Pollen, Wasser und papierartige Masse. Der Deckel ist porös, wodurch die für die Atmung der Larve und dann der Puppe notwendige Luft in die Zelle eindringt. Von diesem Moment an beendet die Larve ihr Wachstum und stellt die Nahrungsaufnahme ein.
Sobald die Zelle verschlossen ist, richtet sich die Larve darin auf, die Darmwände ziehen sich zusammen und die unverdauten Nahrungsreste gelangen, nachdem sie zunächst die dünne Wand des Mitteldarms durchbrochen haben, in die dicke und von dort nach außen. Die Larve lagert ihre Exkremente in einer Ecke der Zelle ab. Danach spinnt die Larve einen Kokon, d. h. sie umflechtet die innere Oberfläche der Zelle. Beim Spinnen eines Kokons nutzt die Larve das Sekret der Spinndrüse, Malpighian-Gefäße und klebrige Sekrete aus den Wänden ihres Körpers. Der Spinnprozess wurde von Velich (1930) ausführlich untersucht und beschrieben.
Während sie den Kokon dreht, macht die Larve schnelle, rhythmische Zitterbewegungen; Ausgehend vom Kopf bewegt sich die Welle durch die Segmente. In 1 Minute macht die Larve bis zu 280 Bewegungen. Gleichzeitig mit dem Zittern macht die Larve mit dem vorderen Ende des Körpers Schaukelbewegungen des Kopfes. Sie haben den Charakter gestrichelter, elliptischer, unregelmäßiger Kurven von oben nach unten oder von rechts nach links. Gleichzeitig fließt aus der Öffnung mit Ventil an der Unterlippe eine transparente Substanz, die die Larve in Form eines dicken oder mehrerer dünner Fäden absondert. Innerhalb von 2 Tagen stärkt die Larve durch Drehen die Wände des Kokons. Beim Spinnen eines Kokons verhielten sich die Larven von Arbeiterbienen, Drohnen und Königinnen ähnlich, der einzige Unterschied bestand in der Form des Kokons. Dies hängt von der Größe der Zelle ab, in der sich die Larve befindet. Bei einer Arbeiterbiene beispielsweise entspricht der Kokon der Form eines sechseckigen Prismas, dessen Boden eine dreiseitige Pyramide ist und dessen Deckel ein exakter Abdruck einer versiegelten Bienenzelle ist. Der Kokon wird nach dem Austritt der jungen Biene nicht entfernt. Aufgrund der Tatsache, dass in einer Zelle Dutzende Generationen dargestellt werden, verringert sich der Durchmesser der Zelle mit der Zeit. Wenn also eine normale Zelle einer frisch gebauten Wabe ein Volumen von 0,282 cm3 und eine Bodendicke von 0,22 mm hat, dann verringert sich nach 20 Generationen von Bienenindividuen die Kubatur der Zelle auf 0,248 cm3 und der Boden verdickt sich auf 1,44 mm .
Die Larve der Arbeiterbiene und der Königin spinnt 2 Tage lang einen Kokon, die Larve der Drohne 3 Tage lang. Anschließend häutet sie sich ein fünftes Mal. Dieser Häutung gehen eine Reihe von Veränderungen im Organismus der Larve voraus. Die Rudimente aller Fortsätze, die sich unter der Nagelhaut befinden, beginnen sich von innen nach außen zu drehen oder sich nach außen zu bewegen und sich in äußere Körperteile zu verwandeln. Diese Organe nehmen nach und nach die für die Puppe charakteristische Form an. Kopf, Mundwerkzeuge und Beine drehen sich von vorne nach hinten-abdominal. Nach dem Spinnen des Kokons richtet sich die Larve auf und gefriert. Es beginnt der Vorbereitungsprozess für das Puppenstadium, der viel Zeit in Anspruch nimmt. Dieser Zeitraum wird als Vorpuppenstadium bezeichnet.
Vorbereitungsstadium. Die Dauer des Vorbereitungsstadiums beträgt für die Arbeitsbiene 3 Tage, für die Königin 2 und für die Drohne 4 Tage. Diese Periode ist durch erhebliche Veränderungen in der Struktur der Larve gekennzeichnet. Die Larvenkutikula wird unter dem Einfluss des speziellen Häutungshormons Ecdyson vollständig von der Puppenkutikula getrennt. Eine Ausnahme bilden die trachealen Verbindungen zwischen Larven- und Puppenspiralen. Der Kopf ist bereits 2/3 so groß wie der Kopf eines Erwachsenen. Die Augen sind vergrößert und ihre Oberfläche wird faltig und faltig. Der Brustkorb ist durch eine deutlich sichtbare Verengung vom Kopf getrennt. Die Flügel beginnen in die Breite zu wachsen, in diesem Stadium sind sie faltig und eng an die Brust gedrückt. Die Mundwerkzeuge verlängern sich und verschmelzen zu einer kompakten Gruppe.
Die Brust- und Bauchsegmente sind entlang einer horizontalen Linie in zwei Hälften geteilt – dorsal und abdominal – und nehmen nach und nach die für einen Erwachsenen charakteristische Struktur an.
Darüber hinaus unterliegen innere Organe dem Verfall. Dieser Vorgang wird Histolyse genannt. Es geht mit dem Eindringen und Einbringen von Blutzellen - Phagozyten - in das Gewebe einher. Die für die diesen Prozessen zugrunde liegenden chemischen Reaktionen notwendigen Energiequellen sind die Reserven an Fetten, Glykogen und anderen Kohlenhydraten im Fettkörper der Larve, der Zucker der Hämolymphe und der Muskeln. Während der Histolyse beginnen Hämozyten als Fresszellen, d. h. als Phagozyten, zu fungieren, und Enzyme wandeln den Fettkörper, die meisten Muskeln der Larve, in ein Nährstoffsubstrat um, das von der Hämolymphe an wachsendes Gewebe abgegeben wird. In den späteren Stadien der Histolyse werden die Phagozyten selbst von den neu entstehenden Geweben aufgenommen. Die Muskulatur erfährt die größten Veränderungen, wodurch die Präpuppe immobilisiert wird.
Im Vorderdarm entsteht ein Honigstruma mit Klappe, der Mitteldarm wird schleifenförmig und nimmt eine gefaltete Struktur an. Im Hinterdarm erscheinen zwei Abschnitte – der Dünndarm und der Mastdarm. Die Anzahl der Tubuli malpighischer Gefäße nimmt zu. Es wird eine teilweise Verschmelzung der Ganglien der Nervenkette beobachtet, wodurch die Puppe sieben Ganglien der Bauchkette anstelle von 11 Ganglien der Larve aufweist. Der Fettkörper wird verkleinert.
Das Vorpuppenstadium endet mit dem Abwerfen der alten Kutikula und dem Herausschlüpfen der Puppe.
Puppenstadium. Die gesamte Körperoberfläche der frisch geschlüpften Puppe ist mit Falten bedeckt, aber nicht faltig. In ihrer Struktur ähnelt die Puppe einer erwachsenen Biene, obwohl sie im Gegensatz zu dieser bewegungslos ist und ihr Körper keine Pigmentierung (weiß) aufweist. Die inneren Lebensprozesse verlangsamen sich jedoch nicht, sondern laufen mit großer Effizienz ab. Die Puppe wird unmittelbar nach Abschluss der letzten Häutung mit einer Nagelhaut bedeckt. Das Puppenstadium ist durch tiefgreifende Veränderungen gekennzeichnet. Die Quelle plastischer Substanzen für die Bildung neuer Organe und Gewebe oder für die Histogenese sind die von Hämozyten transportierten Zerfallsprodukte. Die Spinndrüsen, der Verdauungskanal, der Fettkörper und die Muskeln lösen sich vollständig auf.
Anstelle der verfallenen Organe und Gewebe der Larve werden neue Organe eines erwachsenen Insekts gebildet.
Der Kopf ist viel länger und fast auf die Größe des Erwachsenenkopfes ausgedehnt. Zwischen dem erweiterten Kopf und dem verengten Prothorax befindet sich eine Brücke. Es fehlt jedoch zwischen Brust- und Bauchsegment. Die Puppe hat noch keine Flügel, aber Fühler, Rüssel und Beine sind bereits deutlich ausgeprägt.
Einige der Elemente des Fettkörpers sind während dieser Zeit im Kopf und in der Brust verstreut, andere konzentrieren sich im Bauch. Mit zunehmendem Alter verteilen sich die Bestandteile des Fettkörpers gleichmäßiger im Kopf, in der Brust und im Bauch. Die Struktur des Magens verändert sich.
Das Herz verändert sich kaum: Die Zahl der Kammern verringert sich von 12 auf fünf. Während des gesamten Puppenstadiums hört das Herz nicht auf zu funktionieren, da der Metamorphoseprozess die Bewegung der Hämolymphe erfordert. Die Masse des supraösophagealen Ganglions im Kopf nimmt zu. Dies geschieht aufgrund undifferenzierter Nervenzellen. Darüber hinaus verschmelzen die Nervenganglien im Brustkorb sowie die letzten vier Ganglien am Ende des Bauches.
Die Keimdrüsenrudimente werden nicht zerstört. Ihr Wachstum im Anfangsstadium ist mit dem Zerfall der Zellen des Fettkörpers und der Freisetzung von plastischem Material in Form von Albumiden verbunden. Während ihrer Entwicklung kommen sie mit Teilen der Gonadengänge in Kontakt, die aus dem Ektoderm entspringen.
Einer der Indikatoren für das Innenleben der Puppe ist ihre Atmung.
Das Atmungssystem erfährt während der Histolyse und Histogenese eine Reihe von Veränderungen. So nehmen die ersten und dritten Brustspiralen zu, während die zweiten im Gegenteil abnehmen. Zerstört, Luftröhre. Aus imaginären Rudimenten, die sich an den Trachealstämmen befinden, werden neue Luftröhren sowie Luftsäcke gebildet. 1
Facettenaugen wachsen aus Knospen an den Seiten des Kopfes. Wie oben erwähnt, ist die Puppe zunächst weiß. Während der Entwicklung im Prozess der Bildung verschiedener Organe ändert sich die Farbe der äußeren Haut der Puppe in folgender Reihenfolge: Einen Tag nach der Verpuppung sind die Facettenaugen noch reinweiß, nach 2 Tagen erscheint ein gelblicher Farbton auf dem Am 14. Tag verfärben sie sich rosa und am 16. Tag dunkelviolett.
Die Gelenke der Beine sind zunächst gelblich, ab dem 18. Tag nehmen die Gelenke und Krallen an den Beinen eine gelbbraune Farbe an, die Fersen und Mundwerkzeuge sind etwas heller. Die Truhe wird elfenbeinfarben. Ab dem 19. Tag beginnt sich die Brust leicht zu verdunkeln. Bauch Elfenbein. Die Enden der Beinsegmente sind braun. Eine stärkere Verdunkelung ist am Ende des Kinns und an der Basis des Oberkiefers zu beobachten. Die Augen werden lila. Am 20. Tag ist die Farbe der gesamten Puppe dunkelgrau.
Imago-Bühne. Am Ende des Puppenstadiums wird die Puppenhaut abgeworfen, unter der eine vollständig ausgebildete Biene hervorschlüpft. Mit ihren Kiefern nagt die Biene durch den Deckel der Zelle und gelangt auf die Oberfläche der Wabe. Eine frisch geschlüpfte Biene hat im Vergleich zu einer älteren Biene eine weichere Chitinhülle und außerdem ist ihr Körper dicht mit Haaren bedeckt. Mit der Zeit verlieren Bienen einen Teil ihres Haaransatzes und Chitin wird deutlich härter. Während der Imaginationsperiode treten keine wesentlichen Veränderungen in den äußeren Strukturen von Bienenindividuen auf. "
Stoffwechsel in der postembryonalen Periode. Larve und Puppe durchlaufen intensive Stoffwechselprozesse, wodurch sich die für die Bildung einer erwachsenen Biene notwendigen Nährstoffe ansammeln.
Laut Strauss (1911) ist ein Merkmal des Stoffwechsels von Bienen die Ansammlung erheblicher Mengen an Glykogen als Energie für die Metamorphose. Am Ende des Larvenstadiums erreicht der Glykogengehalt 30 % der Trockenmasse der Larve. Gleichzeitig kommt es zu einer Zunahme der Fettmenge, dem Wachstum von Fettzellen.
Zunächst einmal wird Glukose im Körper der Bienen verbraucht. Sein Gesamtgehalt im Körper der Arbeiterbienenlarven ist doppelt so hoch wie im Körper der Mutterlarven. Wenn der Glukosegehalt in der Hämolymphe unter den zulässigen Wert fällt, beginnen komplexe Kohlenhydrate abgebaut und verwertet zu werden. Die höchste Glukosekonzentration wird in der Mitte des offenen Stadiums beobachtet.
Es ist zu beachten, dass die Aktivität der meisten untersuchten Enzyme bei jungen Larven am höchsten ist. Dann nimmt sie ab und erreicht im Puppenstadium ein Minimum. Gleichzeitig ist die Entwicklung der Enzymaktivität bei Arbeiterbienen bedeutsamer als bei Königinnen. Es besteht die Meinung, dass Königinnen während der postembryonalen Entwicklung im Vergleich zu arbeitenden Individuen einen aeroben Kohlenhydratstoffwechsel haben und die beschleunigte Entwicklung von Königinnen durch die höhere Aktivität einer Reihe von Enzymen erklärt wird. Unterschiede in der Quantität und Qualität der Mitochondrien (Zellorganellen) bei Uteruslarven älter als
- im Vergleich zu denen gleichaltriger Arbeitsbienenlarven. Dementsprechend sind auch einige Unterschiede bei einigen oxidativen Prozessen in Mitochondrien zu beobachten.
Bei der Königinlarve steigt die Sauerstoffverbrauchskurve an, bis die Zelle verschlossen ist. Nach der Versiegelung fällt die Sauerstoffaufnahmekurve jedoch ab (was mit dem Spinnen des Kokons zusammenfällt) und erreicht im anfänglichen Puppenstadium ein Minimum. Der Sauerstoffverbrauch pro Masseneinheit ist bei Königinnen im Vergleich zu Arbeitsbienen und Drohnen am niedrigsten.
Die Menge an Sauerstoff, die pro Individuum bei Drohnen in der postembryonalen Periode aufgenommen wird, ist höher als bei Königinnen und Arbeiterbienen.
Der Atmungskoeffizient, der das Substrat charakterisiert, das während des gesamten Entwicklungsstadiums der Larve einer Arbeitsbiene als Energiequelle verwendet wird, ändert sich und sein Wert übersteigt eins. Wenn der Wert des Atmungskoeffizienten größer als eins ist, zeigt dies die Intensität der Prozesse an, die mit dem Übergang von sauerstoffreichen Verbindungen zu sauerstoffärmeren Verbindungen verbunden sind.
Der Anstieg des Atmungskoeffizienten in der zweiten Phase der Larvenentwicklung fällt mit dem Zeitpunkt des Übergangs zu einer qualitativ anderen Art von Nahrung, hauptsächlich Kohlenhydraten, zusammen. Nach der Versiegelung der Zelle und bis zur Verpuppung der Larven wird der Atmungskoeffizient auf 0,9 eingestellt.
Die Hauptenergiequellen der versiegelten Larve und der Vorpuppe sind in Form von Glykogen abgelagerte Kohlenhydrate und in der zweiten Hälfte des Puppenstadiums Fette.
Im Uterus nimmt im Gegensatz zur Arbeiterbiene der Atmungskoeffizient ab dem 2. Tag der Larvenentwicklung ab, was charakteristisch für den Fettstoffwechsel ist. Am 4.-5. Tag desselben Stadiums entspricht der Atmungskoeffizient fast dem Wert des Atmungskoeffizienten der Arbeiterbienenlarven, was auf die Umwandlung von Kohlenhydraten in Fett hinweist. Im versiegelten Larvenstadium liegt der Atmungsquotient unter eins, was auf den Abbau von Kohlenhydraten hinweist.
Aus dem Vorstehenden können wir schließen, dass der Gasaustausch bei Honigbienen durch eine Reihe insekteninhärenter Muster gekennzeichnet ist. Während der Larvenentwicklung nimmt die Intensität der Atmung, ausgedrückt als verbrauchte Sauerstoffmenge pro Masseneinheit, ab. Allerdings steigt die Menge an Sauerstoff, die pro Larve verbraucht wird, und erreicht vor der Verpuppung ein Maximum. Im Puppenstadium nimmt der Gasaustausch ab und nimmt zum Ausgang des Erwachsenen hin zu. Gleichzeitig gibt es neben Gemeinsamkeiten auch Unterschiede im Atemaustausch der drei Honigbienenformen.
Bei Insekten ist die Entwicklung in zwei Perioden unterteilt – die embryonale und die postembryonale.
Embryonale Entwicklung. Insekteneizellen haben meist eine ovale Form und sind sehr reich an Eigelb, das den gesamten zentralen Teil des Eies ausfüllt. Dementsprechend erfolgt die Zerkleinerung des Eies teilweise und oberflächlich; Dadurch bildet sich eine durchgehende Schicht identischer Zellen, das Blastoderm, das den zentralen Dotter bedeckt. Gleichzeitig verbleibt ein Teil der Kerne in der Dottermasse: Auf ihre Kosten werden in Zukunft spezielle Dotterzellen gebildet. Auf der künftigen Bauchseite des Embryos verdickt sich das Blastoderm. Dieser Bereich wird Keimstreifen genannt; In Zukunft wird auf seine Kosten der Hauptteil des Körpers des Embryos gebildet.
Im Bereich des Keimstreifens beginnen Differenzierungsprozesse. Durch die Fortpflanzung und das Eindringen in die Tiefe der Blastodermzellen entstehen zwei Zellstreifen – Mesodermstreifen, und das Blastoderm selbst wird zum Ektoderm. Später teilen sich die mesodermalen Streifen in paarige Primordien, wodurch eine doppelte Reihe von Zölomsäcken entsteht. Bei höheren Insekten (neg. Diptera) Die Segmentierung mesodermaler Streifen wird unterdrückt und es findet keine Trennung unabhängiger Zölome statt.
Die Frage der Bildung des Endoderms bei Insekten ist noch nicht vollständig geklärt. In vielen Formen scheinen die Vitellinzellen das Rudiment des Endoderms zu sein. Bei Diptera und einigen anderen Insekten werden am vorderen und hinteren Ende des Keimstreifens zwei Gruppen von Zellen getrennt, die auch als Anfänge des Endoderms angesehen werden.
Schon zu Beginn der Keimschichtbildung beginnt der für die Entwicklung von Insekten äußerst charakteristische Prozess der Bildung embryonaler Membranen.
Im einfachsten und am weitesten verbreiteten Fall bildet das Ektoderm entlang der Peripherie des Keimstreifens zunächst eine Rolle und dann eine zweischichtige Falte, die allmählich von allen Seiten zum Keimstreifen wächst, der gleichzeitig etwas eintaucht das Eigelb. Schließlich schließen sich die Ränder der Falte über dem Rudiment des Embryos und lassen zwischen sich und ihm die mit Flüssigkeit gefüllte Fruchthöhle frei, die von der äußeren Umgebung durch eine Doppelmembran getrennt ist. Die äußere Schicht der Membran entsteht durch die Verschmelzung der äußeren Schichten der ektodermalen Falte, die sich über dem Embryo schließt, und wird Serosa genannt, die innere Schicht aus der inneren Schicht derselben Falte, letztere begrenzt direkt die Fruchthöhle und wird Amnion genannt. Serosa und Amnion schützen den Embryo vor Schäden.
In der Zwischenzeit erfährt der Keimstreifen eine Segmentierung und die Segmente sind wie bei anderen isoliert Arthropoden und Ringe, in der Reihenfolge von vorne nach hinten. Im Kopfteil liegen die Augen- und Antennenlappen mit den entsprechenden Rudimenten, das Interkalarsegment, das anschließend einer Reduktion unterzogen wird, und 3 Segmente der Mundglieder. Weiter hinten werden 3 Brustsegmente, die die Rudimente der Gliedmaßen tragen, und 11 Bauchsegmente unterschieden.
Die Differenzierung der inneren Organe besteht in ihren Hauptmerkmalen aus Folgendem. Der Vorderdarm und der Hinterdarm werden durch zwei ektodermale Einstülpungen gebildet. Komplizierter ist die Frage nach der Herkunft des Mitteldarms. Insekten zeichnen sich durch eine erstaunliche Vielfalt an Arten und Quellen ihrer Bildung aus. In einigen Fällen wird es durch endodermale Dotterzellen gebildet, in anderen sind ektodermale Elemente an der Bildung des Mitteldarms beteiligt – spezielle Zellen, die an den blind geschlossenen Enden der Rudimente des vorderen und hinteren Darms lokalisiert sind. Und schließlich wird bei einer Reihe von Arten (parthenogenetische weibliche Blattläuse, einige Schlupfwespen) der Mitteldarm aufgrund des starken Wachstums der Anlage des Vorderdarms, die bis zum hinteren Ende des Körpers reicht, nur vom Ektoderm gebildet. Gleichzeitig degenerieren endodermale Elemente. Malpighische Gefäße entstehen aus dem Ektoderm des Hinterdarms. Das Nervensystem wird in Form einer ventralen Ektodermleiste angelegt, die dann unter die Hautdecke sinkt. Zölomsäcke zerfallen und bilden eine gemischte Körperhöhle – ein Mixocoel. Die gleichzeitig freigesetzten Zellelemente führen zur Entstehung von Muskeln, Herz, Fettkörper und einigen anderen Organen. Primäre Keimzellen werden sehr früh abgelegt, manchmal sogar während der Zeit, in der der Dotter von einer einzigen Blastodermschicht umgeben ist.
Der Embryo wächst, das nährstoffreiche Eigelb wird ständig verbraucht und die Embryonalmembranen platzen, wickeln sich auf der Rückenseite in Form von Falten, die einer Degeneration unterliegen. Der Embryo ist bereit, aus der Eizelle zu schlüpfen, und damit endet die Embryonalentwicklung des Insekts.
Postembryonale Entwicklung. Bei Insekten schlüpft aus dem Ei ein Jungtier oder eine Larve mit voller Segmentzahl. Die einzigen Ausnahmen sind primitive Kryptomaxillarien, die zu neg gehören. Protura, wobei Jugendliche sich von Erwachsenen durch das Fehlen von drei hinteren Abdomensegmenten unterscheiden. Letztere werden nacheinander nach den ersten Häutungen aus der hinteren Wachstumszone gebildet. Somit ist eine Entwicklung ohne Anamorphose typisch für Insekten, und nur bei Protur findet sich eine Restanamorphose.
Die postembryonale Entwicklung von Insekten zeichnet sich durch eine erhebliche Vielfalt aus, die maßgeblich durch die biologischen Eigenschaften der Vertreter dieser Tiergruppe bestimmt wird.
In den primitivsten Formen (subcl. Entognatha) Das Wachstum und die Entwicklung eines jungen Organismus gehen nicht mit wesentlichen strukturellen Veränderungen einher. Jungtiere ähneln in allen Hauptmerkmalen dem Organismus der Mutter, d. h. das Larvenstadium und die Metamorphose fehlen. Im Wesentlichen handelt es sich in diesem Fall um eine direkte Entwicklung. Geflügelte Insekten, die evolutionär höher stehen, weisen dagegen eine Metamorphose auf und werden je nach Art der Transformation in hemimetabolische – mit unvollständiger Transformation und holometabolische – mit vollständiger Metamorphose unterteilt. Eine unvollständige Metamorphose ist charakteristisch für niedriger organisierte Gruppen geflügelter Insekten: Orthoptera, Kakerlaken, Käfer, Eintagsfliegen, Libellen. Bei unvollständiger Transformation ähnelt die aus dem Ei schlüpfende Larve im allgemeinen Erscheinungsbild bereits in vielerlei Hinsicht dem erwachsenen Organismus und unterscheidet sich von ihm hauptsächlich durch den embryonalen Zustand der Flügel, die Unterentwicklung sekundärer Geschlechtsmerkmale und häufig das Vorhandensein einiger vorläufiger Merkmale , reine Larvenorgane: zum Beispiel Trachealkiemen bei Eintagsfliegenlarven usw. Flügel haben zunächst nur die Form kaum sichtbarer äußerer Rudimente. Die Glättung des Unterschieds zwischen der Larve und dem erwachsenen Organismus erfolgt allmählich, begleitet von einer Reihe von Häutungen.
Mit jeder weiteren Häutung ähnelt die Larve immer mehr dem Erwachsenenstadium von Insekten oder Erwachsenen (Imago), die Flügelrudimente nehmen zu usw.
Bei einigen primitiven Insekten ist die Anzahl der Häutungen sehr groß und kann 25–30 (Eintagsfliegen) erreichen. Vertreter anderer Abteilungen haben weniger Linien (4-5).

Bei holometabolischen Insekten ist die Entwicklung anders. Die Larve unterscheidet sich stark vom erwachsenen Tier: Sie hat überhaupt keine äußeren Flügelansätze, sie hat oft eine andere Anzahl von Gliedmaßen usw. Die Larvenmerkmale bleiben während einer Reihe von Häutungen, d. h. während der gesamten Larvenperiode, erhalten Leben. Die Larven holometabolischer Insekten sind äußerst vielfältig. Die Larve unterscheidet sich von der Imago durch eine Reihe primitiver Merkmale: eine größere Homonomie der Dissektion, das Fehlen von Flügeln, eine schwache Entwicklung der Sinnesorgane (es gibt keine Facettenaugen, sondern nur Augenflecken, Antennen sind sehr klein, usw.), und in vielen Fällen auch die Vorrichtung und Anzahl der Gliedmaßen. Sehr oft sind die Mundglieder im Larvenalter nach einem Typ angeordnet (Nagen bei Schmetterlingsraupen), bei Erwachsenen anders (Saugmund bei erwachsenen Schmetterlingen).
Wie bereits erwähnt, bleibt die für die Larve charakteristische Struktur während der gesamten Larvenperiode erhalten: Die Transformation erfolgt nicht schrittweise, sondern die für die Metamorphose notwendigen Veränderungen werden auf den Zeitpunkt der letzten Larvenhäutung verschoben. In dieser Zeit finden so grundlegende Veränderungen im Organismus der Larve statt, dass die Aufrechterhaltung der gewohnten physiologischen Funktionen, insbesondere Ernährung und Bewegung, schwierig, ja sogar unmöglich wird. Dies spiegelt sich darin wider, dass die Larve nach der letzten Häutung in einen Ruhezustand übergeht – sie wird zur Puppe. Die Puppe stellt das charakteristischste Stadium der holometabolischen Transformation dar. Je vollständiger der Ruhezustand ist, desto besser kommt die vollständige Transformation zum Ausdruck.
Die individuelle Entwicklung eines Insekts oder die Ontogenese kann in zwei Perioden unterteilt werden – die Entwicklung innerhalb der Eizelle oder die embryonale Entwicklung und die Entwicklung nach dem Verlassen der Eizelle oder die postembryonale.
Die meisten Insekten legen Eier. Ein Insektenei ist eine Zelle, die einen Zellkern, Protoplasma, Deutoplasma oder Eigelb enthält, die für die Ernährung und Entwicklung des Embryos notwendig sind. Darüber hinaus enthalten Insekteneier oft symbiotische Mikroorganismen, die von der Mutter über die Eierstöcke (d. h. transovarial) gewonnen werden. Das Ei ist außen mit einem Chorion – einer Schale – bedeckt. Das Chorion weist manchmal eine ausgeprägte Mikroskulptur auf, die als zuverlässiges Zeichen für die Unterscheidung von Insektengattungen und sogar Insektenarten entsprechend der Eiphase dient. Unter dem Chorion liegt die wahre oder Dottermembran des Eies. Mit der Bildung des Chorions entsteht eine Öffnung zur Mikropyle, die dem Durchgang des Spermatozoons während der Befruchtung dient. Das Ei hat ein vorderes und ein hinteres Ende, die der Position des Embryos entsprechen. Im Eierstock ist der vordere Pol des Eies auf den Kopf der Mutter gerichtet bzw. befinden sich die dorsalen und ventralen Seiten des Eies.
Die Größe und das Aussehen der Eier sind unterschiedlich. Die Größe beträgt 0,02 – 0,03 mm bei Blattläusen und Thripsen, 8 – 10 mm bei Heuschrecken. Die Eier sind bei Blattläusen glatt, bei Schaufeln gerippt, weiß, manchmal mit einem Deckel versehen und mit Falten bedeckt.
Eiform. Oval - Käfer, Schmetterlinge - länglich, halbkugelförmig - Schaufeln, flaschenförmig - tagaktive Schmetterlinge, tonnenförmig - Käfer, haben einen Stiel - Florfliegen, Saugnäpfe. Eiablage: einzeln, in Gruppen, offen oder im Substrat, kann geschützt sein und mit Gonadensekreten bedeckt sein.
Die Embryonalentwicklung beginnt mit der Fragmentierung des Zellkerns und der Bewegung der daraus resultierenden Tochterkerne mit kleinen Protoplasmabereichen an die Peripherie der Eizelle. Hier bildet sich aus der Masse der Tochterkerne eine durchgehende Zellschicht – das Blastoderm. Später differenziert sich das Blastoderm in die Keim- und die extraembryonale Zone: Die Zellen der letzteren sind nicht an der Bildung des Embryos beteiligt, während die Zellen der Keimzone beginnen, sich intensiver zu teilen und auf der ventralen Seite ein Embryonalband zu bilden des Eies, das dann hervorsteht und Ektoderm und Mesoderm entstehen.
Die Entwicklung des Embryos wird von der Blastokinese begleitet: Embryonalmembranen werden gebildet, die Kopfsegmente, Brust- und Bauchsegmente werden getrennt. Mit der Bildung des Mesoderms wird der Grundstein für die innere Struktur gelegt. Aus dem Ektoderm gehen alle äußeren Hüllen hervor – dann Mund und Anus, der vordere und hintere Darm. Aus dem Mesoderm entstehen das Muskelsystem, der Fettkörper, das Rückengefäß und die Scheide der Geschlechtsdrüsen.
Der Embryo beginnt sich zu bewegen, fängt Luft und Flüssigkeit aus dem Ei ein, das Chorion bricht und der Embryo, der sich in eine Larve verwandelt hat, kommt heraus.
2. Die Fortpflanzung von Insekten weist eine Reihe von Merkmalen auf, die einen wesentlichen Aspekt ihrer Biologie darstellen und berücksichtigt werden müssen. Zu diesen Merkmalen gehören Fortpflanzungsmethoden, zusätzliche Ernährung, Begegnung der Geschlechter und Befruchtung sowie sexuelle Produktivität.
Reproduktionsmethoden .
Die Fortpflanzung geht bei den meisten Insekten mit der Paarung und Befruchtung einher, d.h. mit der Beteiligung zweier Geschlechter verbunden – daher wird es bisexuell (bisexuell) gamogenetisch genannt. Die meisten Insekten legen Eier, es gibt jedoch auch andere Fortpflanzungsmethoden: Lebendgeburt, Parthenogenese, Pedogenese, Polyembryonie. Bei der Lebendgeburt wird die Embryonalentwicklung im Körper der Mutter abgeschlossen. Anstatt Eier zu legen, werden Larven (Blattläuse, Bremsen, blutsaugende Fliegen) zur Welt gebracht. Die Larven schlüpfen in den Eiröhren oder beim Durchgang durch die Eileiter.
Parthenogenese – jungfräuliche Fortpflanzung ist durch das Fehlen einer Befruchtung gekennzeichnet und wird sowohl bei eierlegenden als auch lebendgebärenden Insekten beobachtet.
Formen der Parthenogenese:
1. Arrhenotokie- Aus unbefruchteten Eiern entwickeln sich nur Männchen. 2) thelytokia - nur Weibchen entwickeln sich aus unbefruchteten Eiern; 3) Amphitokie- Männchen und Weibchen entwickeln sich aus unbefruchteten Eiern.
Parthenogenese kann fakultativ, permanent und zyklisch sein.
Fakultative Parthenogenese oder spadisch manifestiert sich uneinheitlich, tritt unter äußeren Einflüssen oder bei einem bestimmten physiologischen Zustand des eierlegenden Weibchens auf (Zigeunermotte, Pappelschwärmer).
Die ständige Parthenogenese vom Arrhenotokie-Typ ist typisch für soziale Insekten. Bei Bienen entwickeln sich aus unbefruchteten Eiern immer Männchen und aus befruchteten Eiern Weibchen. Eine permanente Parthenogenese vom Typ Thelytoky wird bei Insekten beobachtet, die keine koloniale Lebensweise haben. So fehlen bei vielen Würmern, Thripsen, Blattwespen und einigen Reitern Männchen, die Fortpflanzung erfolgt immer durch Parthenogenese und aus unbefruchteten Eiern schlüpfen nur Weibchen.
Parthenogenese spielt im Leben von Insekten eine sehr wichtige Rolle. Dank der Parthenogenese und ihren Modifikationen erhöht sich das Fortpflanzungspotenzial der Art dramatisch: Durch die parthenogenetische Fortpflanzung werden die Überlebenschancen der Art unter widrigen Bedingungen und während der Besiedlung deutlich erhöht.
3. Nach dem Verlassen der Eizelle beginnt die postembryonale Entwicklung des Insekts. Diese Periode ist kein einfaches Wachstum und eine Zunahme der Körpergröße, sondern ist durch den Übergang des Körpers von einer Phase in eine andere gekennzeichnet. Diese Art der individuellen Entwicklung nennt man Metamorphose oder Entwicklung mit Transformation.
Das Wesen der Metamorphose liegt in der Tatsache, dass ein sich entwickelndes Individuum im Laufe seines Lebens eine erhebliche Umstrukturierung seiner morphologischen Organisation und seiner biologischen Merkmale durchläuft. In dieser Hinsicht gibt es eine Differenzierung der postembryonalen Entwicklung in zwei Phasen – Larven- und Erwachsenenphase. In der Larvenphase findet das Wachstum und die Entwicklung der Individuen statt, in der Imagophase die Fortpflanzung und Ansiedlung. In anderen Fällen gibt es zwischen diesen Phasen eine Zwischenphase – die Puppe. Es gibt zwei Haupttypen der Metamorphose: unvollständige und vollständige Transformation. unvollständige Konvertierung oder Hemimetamorphose 3 Phasen – Ei, Larve, Adult. Die Larven ähneln erwachsenen Tieren (Komplexaugen, Mundwerkzeuge, Flügelrudimente, ähnliche Lebensweise), imagoartig. Mit vollständiger (Holometamorphose) 4 Phasen. Die Larven sehen überhaupt nicht wie Erwachsene aus, es gibt keine komplexen Augen und Flügel, sie leben unter unterschiedlichen Bedingungen.
Manche haben Bauchbeine, einen anderen Mundapparat, andere haben Seiden- oder Spinnendrüsen.
Modifikationen der unvollständigen Transformation: a) Hypomorphose – eine vereinfachte unvollständige Transformation und ist charakteristisch für geflügelte Insekten, die Flügel verloren haben (sekundäre flügellose Läuse, Läuse, flügellose Vertreter von Heuschrecken, Heuschrecken, Grillen, Kakerlaken, Stabheuschrecken, Heufresser, Wanzen); b) Hypermorphose – ein komplizierter unvollständiger Zyklus (Aleurodiden, Thripse, Kokzidenmännchen). Das Erscheinen der Larven im Ruhezustand am Ende der Phase.
Anamorphose- Larven ähneln erwachsenen Tieren, haben aber eine geringere Anzahl an Bauchsegmenten; während sich die Larve entwickelt, wachsen zusätzliche Segmente an der Oberseite des Bauches, die volle Anzahl erreicht sie jedoch erst im Erwachsenenstadium.
Protomorphose– Häutung im erwachsenen Zustand, eine gewisse Ähnlichkeit der Larve mit der erwachsenen Phase, aber das Fehlen einer Teilung der Larve in Brustkorb und Hinterleib (Podura, zweischwänzig, borstenschwänzig).
Hypermetamorphose- Komplikation der vollständigen Transformation (das Vorhandensein von 2 Larvenformen und manchmal Puppen (Blasen)).
4. Die Larvenphase beginnt unmittelbar nach dem Schlüpfen aus dem Ei. Anfangs hell, nach dem Schlüpfen ist die Larve farblos, hat weiche Hüllen. Bei offen lebenden Insekten verdunkelt sich die Larve schnell und verhärtet sich. Nach der Verdauung der Reste des embryonalen Dotters und der Ausscheidung von Exkrementen entstehen Fressreize. Die Larve durchläuft ein Paar verbesserter Ernährung, Wachstum und Entwicklung. Ihr Wachstum und ihre Entwicklung gehen mit periodischen Häutungen und dem Ablösen der Hautkutikula einher – durch Häutungen nimmt der Körper zu und es kommt zu großen Veränderungen. Die Anzahl der Glieder variiert zwischen 3 (Fliegen), 4–5 (Käfer), 25–30 (Padenki).
Nach jeder Häutung tritt die Larve in das nächste Stadium oder Alter ein, daher teilen sich die Häutungen das Alter der Larven. Entsprechend der Anzahl der Häutungen ergibt sich auch die Anzahl der Larvenstadien. Die Bestimmung des Häutungsalters (notwendig für die Planung des Kampfzeitpunkts) erfolgt anhand folgender Merkmale: Antennen bei Heuschrecken, Vergrößerung der Körpergröße (Breite der Kopfkapsel) bei Schmetterlingsraupen und Käferlarven. Es gibt zwei Arten von Larven: erwachsene (primäre) und sekundäre.
Najaden sind Larven mit Kiemen, die im Wasser leben (Libellen, Padyonki, Steinfliegen).
Campodeoid – beweglich, dunkel gefärbt mit dichter Haut und mit 3 Paar Brustbeinen, einem gut isolierten prognathen Kopf (Laufkäfer, Schwimmer, Florfliegen).
Wurmartig – sesshaft, hell, ohne Bauchbeine.
Raupen haben eine gut getrennte Kopfhaut, 3 Paar Brustbeine und 2-8 Paar Bauchbeine.
Puppenphase. Diese Entwicklungsphase der Metamorphose ist nur für Insekten mit einem vollständigen Zyklus charakteristisch. Ein Merkmal der Puppe ist die Unfähigkeit, Nahrung aufzunehmen und sehr oft unbeweglich zu bleiben. Es lebt von den von der Larve angesammelten Reserven und wird oft als Ruhephase angesehen.
Äußerlich sieht die Puppe nicht wie ein Erwachsener aus, weist jedoch eine Reihe von Anzeichen der Erwachsenenphase auf: äußere Rudimente von Flügeln, Beinen, Antennen, Facettenaugen usw. Histolyse und Histogenese finden in der Puppenphase statt.
Histolyse- der Zerfall der inneren Organe der Larve, der mit dem Eindringen und Eindringen von Blutzellen - Hämozyten - in das Gewebe einhergeht. Hämozyten fungieren als Zellfresser, d.h. Fresszellen, deren erhöhte Aktivität zur Zerstörung und Absorption von Gewebematerial führt. Die Histolyse erfasst das Muskelsystem, sodass die Vorpuppe unbeweglich wird, das Verdauungssystem beeinträchtigt, jedoch nicht das Nerven- und Fortpflanzungssystem sowie das Rückengefäß beeinträchtigt.
Histogenese- der Prozess der Schaffung von Geweben und Organen des imaginären Lebens. Die Quelle für die Bildung dieser neuen Gewebe und Organe sind die Produkte der Histolyse. Die Histogenese umfasst das Muskel- und Verdauungssystem und ordnet sie für neue, imaginäre Funktionen neu. Die Muskulatur wird für neue Bewegungsformen beim Fliegen umgebaut, das Verdauungssystem für neue Nahrungsarten.
Bei der Histogenese spielen imaginäre Rudimente die Hauptrolle – Gruppen von Unterhautzellen, aus denen bestimmte Gewebe und Organe entstehen.
Das Häutungshormon ist Exidon, das für die normale Entwicklung der Larven notwendig ist.
Es gibt Folgendes Puppentypen:
offene Puppen- freie, nur an den Körper gedrückte Imaginalfortsätze (Antennen, Beine, Flügel), charakteristisch für Käfer.
Puppen mit beweglichen Mandibeln – verwenden bewegliche Oberkiefer, um den Kokon zu verlassen, und können sich selbst bewegen (Retikulat, Köcherfliegen, Zahnmotten). Puppen mit unbeweglichen Mandibeln können diese beim Austreten aus dem Kokon nicht nutzen (Käfer, Hautflügler, Fanopteren, viele Zweiflügler).
Versteckte Puppen bedeckt mit verhärteter, sich nicht ablösender Larvenhaut, die die Rolle einer Muschel oder eines falschen Kokons spielt - Puparia (Fliegen).
bedeckt- haben imaginäre Anhängsel, die eng mit dem Körper verbunden und verlötet sind, da die Larve während der letzten Häutung ein Geheimnis absondert, das, wenn es ausgehärtet ist, die Puppe mit einer harten Schale bedeckt (Schmetterlinge, Marienkäfer).
Einige Larven umgeben sich vor der Verpuppung mit einem Kokon (Seide, Spinnennetz). Manchmal sind Pflanzenstängel, Erdwiege oder offene Verpuppung der Ort der Verpuppung.
Imago-Phase.
Die aus der Puppe schlüpfenden Insekten weisen Anzeichen eines Erwachsenen auf, die Flügel bleiben jedoch zunächst gefaltet. Dann breiten sie ihre Flügel aus, die Haut verdickt sich und nimmt die Farbe eines voll ausgebildeten erwachsenen Insekts an. Im Erwachsenenalter häuten sich Insekten nicht und sind nicht wachstumsfähig. Die Ausnahmen sind Padyonki, Narren. Die biologische Funktion der Erwachsenenphase besteht darin, sich auszubreiten und zu reproduzieren. Dies ist bereits eine Funktion des Artenlebens und zielt auf die Erhaltung der Art ab. Dank der Flügel erhöht sich die Fähigkeit erwachsener Insekten, sich niederzulassen, erheblich und die Fortpflanzung ermöglicht es Ihnen, Nachkommen an einem neuen Ort zurückzulassen.
Die Ausbreitung erwachsener Insekten erfolgt sowohl durch aktiven als auch durch passiven Flug. Aktive Flüge sind in der Regel charakteristisch für große Insekten und werden bei einer Reihe von Libellen-, Heuschrecken-, Schmetterlings- und Käferarten beobachtet und sind weit verbreitet. Passivflüge sind charakteristisch für Blattläuse und Fliegen.
Der Übergang zum Erwachsenen geht einher mit äußeren Veränderungen der Körperfarbe, einer Vergrößerung des Bauches beim Weibchen, aufgrund der Entwicklung von mit Eiern gefüllten Eierstöcken – bei befruchteten Weibchen von Termiten und Ameisen – fallenden Flügeln, bei unreifen Heuschrecken – rosa , reif - leuchtend gelb.
Geszwischen Männchen und Weibchen in einer Reihe äußerer, sekundärer Geschlechtsmerkmale – in Form und Größe der Fühler, Körpergröße, verschiedenen Strukturdetails (Nashornkäfer, Hirschkäfer). Männer sind mobiler und führen einen offeneren Lebensstil.
Polymorphismus – die Existenz von Insekten, äußerlich unterschiedlichen Formen derselben Art – (Ameisen, Bienen, Termiten, Männchen, Weibchen, Arbeiter, Soldaten). Sexueller Polymorphismus wird innerhalb der Familie kontrolliert und hängt nicht vom Einfluss äußerer Faktoren ab.
Ökologischer Polymorphismus entsteht unter dem Einfluss der äußeren Umgebung (langflügelige, kurzflügelige, flügellose Käfer).