The embryonic development of insects typically begins with superficial crushing of the egg, which is associated with the rich content of the last nutritious yolk. The bastoderma formed on the surface of the egg is thicker in some part of the surface; this thickening gives rise to the germinal streak and defines the ventral side of the embryo.
The annular fold formed along the edge of the germinal strip, overgrowing and growing together above the germinal strip, gives two layers of germinal membranes - the inner (amnion) and outer (serosa). From the cell layer of the germinal strip on the side facing the yolk,the primary endoderm is separated - the common germ of the endoderm and mesoderm. Subsequently, the germinal streak is segmented.
The postembryonic development of insects is very diverse due to the variety of conditions for the existence of insect juveniles and the degree of its adaptation to them. Two main types of postembryonic development can be distinguished: incomplete transformation, hemimetabolism, and complete transformation, holometabolism. Holometabolism is characterized by profound changes in juveniles, as a result of which the juveniles differ sharply in structure from the adult insect (adult). Therefore, the transition from the state of juveniles, i.e., from the larval phase to the imaginal phase, requires a radical restructuring of the organs; this is achieved with the participation of an intermediate phase between the larval and imaginal phases, during which the juveniles remain dormant; this intermediate phase is pupal. During the pupal state, the internal organs of the insect either undergo more or less restructuring, or undergo complete destruction (histolisis) and are rebuilt; in the latter case, the formation of the organ anew occurs due to the so-called imaginal discs, consisting of cells of an embryonic nature (histoblasts). Thus, the pupal period is a period of active histolysis and histogenesis (tissue neoplasms). Outwardly, the pupa represents the imaginal phase with its characteristic organs, which, however, remained underdeveloped.
The development in higher orders of insects is holometabolic in beetles, lepidoptera, caddisflies, fleas, hymenoptera and dipterans.
In the case of incomplete transformation - hemimetabolism - there is no such profound difference in the structure of the young and adult phases: the juvenile now more, then somewhat less resembles the imaginal phase in structure. Finally, in cases where the transition from the young phase to the adult is reduced only to growth and shedding ( in connection with the growth) of the old cover (exuvia), they speak of amebolism, that is, the absence of transformation; an example of amebolism is the development of Collembola among the Apterygota. However, usually with successive instars, which are separated from each other by molts, that is, by the shedding of the cover of the previous age, changes in the shape of the organs also occur: for example, the length of the antennae and cerci increases relatively, the number of segments increases; some organs may reappear, such as the external genital appendages. Such is the development in some Apterygota, such as Thysanura and Diplura, and in most of the hemimetabolic Pterygota. In some Pterygota developing with hemimetabolism (dragonflies, mayflies; stoneflies and parts of arthropods), development is complicated due to the fact that juveniles live in other conditions than adults; thus, juveniles of dragonflies, mayflies, and stoneflies develop in water bodies and, in connection with this, acquire organs of water respiration - gills, which are an example of larval (provisional) organs. The conditions of existence are also reflected in the development of other organs of the larva, which adapt to the environment, for example, the transformation of the front legs of the juvenile cicada into a digging organ in connection with its underground life.
Insect development
The individual development of insects (ontogeny) consists of embryonic development occurring in the egg phase, and postembryonic development - after the larva leaves the egg until reaching the adult phase - the imago.
Embryonic development. Insect eggs vary in shape due to adaptations to the environment in which they develop. For example, in beetles, eggs are predominantly oval and develop more often in a closed substrate; in bedbugs - barrel-shaped, attached to the substrate; butterflies - turret-shaped or bottle-shaped; in lacewings (golden eyes) eggs with a stalk. Eggs are often laid in groups. Egg clutches may be open or closed. An example of open laying is the eggs of the Colorado potato beetle, glued by the female to the underside of potato leaves. Closed clutches include egg-pods of acridoids formed from soil particles cemented by excretions of the adnexal glands of females. Cockroaches lay their eggs in ootheca - egg capsules that form in the female genital tract.
Insect eggs are covered on the outside with a shell - chorion, which protects them from drying out (Fig. 337). On the surface of the shell there is a micropyle - a small hole with a complex "plug" with a tubule inside for the penetration of spermatozoa during fertilization. Under the chorion is a thin yolk membrane, and under it is a dense layer of cytoplasm. The central part of the cytoplasm is filled with yolk. The cytoplasm contains the nucleus and polar bodies.
Crushing is superficial. Initially, the nucleus divides many times, the daughter nuclei with sections of the cytoplasm migrate to the periphery of the egg, are covered with a membrane and a surface layer of cells is formed - the blastoderm, and the yolk remains in the center of the egg. On the ventral surface of the blastoderm, the cells are higher and form a thickening - the germinal strip. This stage of insect embryonic development corresponds to the blastula.
Cell division of the germinal strip leads to the development of the embryo. The germinal strip gradually sinks, forming the ventral groove.
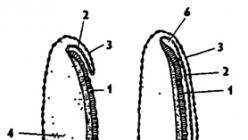
Rice. 337. The structure of an insect egg (from Bei-Bienko): 1 - micropyle, 2 - chorion, 3 - yolk membrane, 4 - nucleus, 5 - polar bodies, 6 - yolk

The folds of the blastoderm above the furrow close, and germinal membranes are formed: serosa and amnion (Fig. 338). Here, convergence with higher vertebrates, which also have similar shells, is manifested. Due to the amniotic cavity formed, the embryo is in a suspended state inside the egg, which reliably protects it from mechanical damage. In addition, the fluid that fills the amniotic cavity facilitates the metabolic processes of the embryo.
The germinal strip further differentiates into two layers: the lower - ectoderm and the upper - endomesoderm. Entomesoderm in different species of insects can be formed in different ways: by invagination or cell immigration.
At the next stage of development, the ectodermal layer of the strip begins to bend upwards on the sides, and then closes on the back, forming a closed wall of the embryo. When the walls of the body close on the back, part of the yolk and yolk cells enters the body of the embryo. Simultaneously with the formation of the walls of the body of the embryo, two groups of cells are separated in the entomesoderm at the anterior and posterior ends of the body. These are the two rudiments of the midgut. Subsequently, the anterior and posterior sections of the midgut begin to form from these two primordia, which then merge. At the same time, deep invaginations of the ectoderm form at the anterior and posterior ends of the body of the embryo, from which the anterior and posterior sections of the intestine are formed. Then all three sections are connected, forming a through intestinal tube.
The mesodermal streak breaks up into paired metameric rudiments of coelomic sacs. But later they disintegrate, and the muscles of the embryo, the somatic sheet of the coelomic epithelium, the heart, the fat body and the gonads are formed from the mesoderm. The visceral layer of the coelomic epithelium is not formed in insects, and the body cavity becomes mixed - the mixocoel. Coelomic rudiments merge with the primary body cavity.
Later, the nervous system and the tracheal system are formed from the ectoderm. Malpighian vessels form from the walls of the posterior intestine.
In the process of development, the insect embryo undergoes segmentation, which first manifests itself in the anterior part, and then in the posterior part of the body. In the head section, an acron with eye, labial and

antennal lobes, an intercalary segment, and three jaw segments. Then three thoracic and ten abdominal segments and an anal lobe are formed.
In many insects, the embryo passes through three stages, characterized by a different composition of the limb primordia: protopod, polypod, and oligopod (Fig. 339).
The embryonic development of insects is characterized by the phenomenon of blastokinesis. This is a change in the position of the body of the embryo in the egg, in which the yolk reserves are most fully used.
Two types of blastokinesis for insects are described by A. G. Sharov. In insects with incomplete transformation, at first the embryo is located with its back up and head towards the anterior end of the egg, and then, when the amniotic cavity is formed, the embryo turns over with the ventral side up, and the head, respectively, is in the back of the egg.
Blastokinesis occurs differently in most insects with complete transformation and in orthoptera, in which the embryo sinks into the yolk without changing the position of the body in the egg.
In the embryonic development of insects, adaptations to life on land are manifested: protective membranes (chorion, serosa, amnion), a supply of nutrients (a lot of yolk), an amniotic cavity filled with liquid.
Before hatching, the formed insect larva swallows fluid from the amniotic cavity, due to which the body turgor increases. The larva breaks through the chorion with its head, which often has egg teeth or a spike.
Postembryonic development. During the period of postembryonic development of insects, after hatching from the egg, the growth of a young animal occurs through successive molts and the passage of qualitatively different phases of development. During ontogenesis, or individual development, insects molt from 3-4 to 30 times. On average, the number of links is 5-6. The interval between molts is called the stage, and the state of development is called age. Morphological changes during development from larva to adult are called metamorphosis. In all insects, except for the lower wingless forms, after reaching the adult state, growth and molting of adults cease. Therefore, for example, variations in the size of beetles of the same species cannot be attributed to different age groups, but should be considered only a manifestation of individual variability.
There are three main types of postembryonic development of insects: 1) direct development without metamorphosis - ametabolism, or protometabolism; 2) development with incomplete transformation, or with gradual metamorphosis, - hemimetabolism; 3) development with complete transformation, i.e. with a pronounced metamorphosis - holometabolism.
Ametabolism, or direct development, is observed only in primary wingless insects from the bristle-tailed order (Thysanura), which include the common silverfish (Lepisma). The same type of development is observed in Entognatha: springtail (Collembola) and two-tailed (Diplura).
With ametabolism, a larva, similar to an adult, emerges from the egg. The differences concern only the size, proportions of the body and the degree of development of the gonads. Unlike winged insects, they continue to molt even in the imaginal state.
Hemimetabolism- incomplete transformation, or development with gradual metamorphosis. It is characteristic of many winged insects, for example, cockroaches, grasshoppers, locusts, bedbugs, cicadas, etc.
With hemimetabolism, a larva emerges from the egg, similar to adults, but with rudimentary wings and underdeveloped gonads. Such adult-like larvae with rudiments of wings are called nymphs. This name is borrowed from ancient Greek mythology and refers to divine winged beings in the form of girls. Insect nymphs molt several times, and with each molt, their wing rudiments increase. An older nymph molts, and a winged imago emerges from it. Figure 340 shows the phases of locust development (eggs, nymphs of 1st-5th instars and adults) as an example of incomplete transformation. This typical incomplete transformation is called hemimetamorphosis.
Among insects with incomplete metamorphosis, there are cases of development when nymphs noticeably differ from adults in the presence of special larval adaptations - provisional organs. Such development is observed in dragonflies, mayflies, stoneflies. The nymphs of these insects live in water, and therefore they are called

Rice. 340. Development with incomplete metamorphosis in the locust Locusta migratoria (according to Kholodkovsky): 1 - prothorax, 2 - mesothorax with wing buds, 3 - metathorax with wing buds
naiads (water nymphs). They have provisional organs such as tracheal gills, which disappear in land adults. And dragonfly larvae still have a "mask" - a modified lower lip that serves to grasp prey.
Holometabolism- complete transformation. The developmental phases in holometaboly are: egg - larva - pupa - imago (Fig. 341). This development is typical for beetles, butterflies, dipterans, hymenoptera, caddisflies and lacewings.

Rice. 341. Development with complete transformation in the silkworm Bombyx top (according to Leines): A - male, B - female, C - caterpillar, D - cocoon, E - cocoon pupa
Insect larvae with complete metamorphosis are not similar to adults and often differ ecologically. So, for example, the larvae of the May beetles live in the soil, and the adults live on trees. The larvae of many flies develop in the soil, rotting substrate, and adults fly and visit flowers, feeding on nectar. The larvae of such insects molt several times and then turn into a pupa. At the pupal phase, histolysis occurs - the destruction of larval organs and histogenesis - the formation of the organization of an adult insect. A winged insect emerges from the pupa - an imago.
Thus, the following types of postembryonic development are observed in insects: ametabolism, or protomorphosis (egg - larva (similar to imago) - imago); hemimetabolia - incomplete transformation (egg - nymph - imago): hemimetamorphosis - a typical variant, hypomorphosis - reduced metamorphosis, hypermorphosis - increased metamorphosis; holometaboly - complete transformation (egg - larva - pupa - adult): holometamorphosis - a typical variant, hypermetamorphosis - with several types of larvae.
Types of insect larvae with complete metamorphosis. Larvae of holometabolic insects have a more simplified structure compared to adults. They do not have compound eyes, rudiments of wings; the mouth apparatus is of a gnawing type, the antennae and legs are short. According to the development of the limbs, four types of larvae are distinguished: protopods, oligopods, polypods, and apods(Fig. 342). Protopod larvae are characteristic of bees, wasps. They have only the beginnings of pectoral legs. These larvae are inactive, develop in combs with care from workers. Oligopod larvae are more common than others; they are characterized by the normal development of three pairs of walking legs. Oligopods include larvae of beetles, lacewings. Polypod larvae, or caterpillars, have, in addition to three pairs of thoracic legs, several more pairs of false legs on the abdomen. The ventral crura represent protrusions of the abdominal
body walls and bear hooks and spines on the sole. Caterpillars are characteristic of butterflies and sawflies. Apodic, or legless, larvae are observed in the Diptera order, as well as in some beetles (larvae of barbels, borers), butterflies.

Rice. 342. Insect larvae with complete metamorphosis (from Barnes): A - protopod, B, C - oligopod, D - polypod, E, F, G - apod
According to the methods of movement, insect larvae with complete metamorphosis are divided into campodeoid with a long, flexible body, running legs and feeling cerci, erucoid with a fleshy, slightly curved body with or without limbs, wire- with a rigid body, round in diameter, with supporting cerci - urogomphs and worm-like- legless.
Campodeoid larvae are characteristic of many predatory beetles - ground beetles, rove beetles. They move through holes in the soil. A typical erucoid larva is the larva of the May beetle, dung beetle, bronze. These are burrowing larvae. Wireworms are characteristic of click beetles and dark beetles, the larvae of which actively make passages in the soil. Numerous worm-like larvae. They move in the soil, plant tissues. These include not only Diptera larvae, but also some beetles, butterflies, sawflies, developing, for example, in plant tissues.
types of pupae. Pupae are free, covered and hidden (Fig. 343). In free pupae, the rudiments of wings and limbs are clearly visible and freely separated from the body, for example, in beetles. In covered pupae, all rudiments adhere tightly to the body, for example, in butterflies. The integuments of free pupae are thin, soft, while those of the covered pupae are strongly sclerotized.

Rice. 343. Types of pupae in insects (from Weber): A - free beetle, B - covered butterfly, C - hidden fly; 1 - antenna, 2 - rudiments of wings, 3 - leg, 4 - spiracles
larval skin, which forms a false cocoon - puparia. Inside the puparium is an open pupa. Therefore, the hidden chrysalis is only a variant of the free one. Puparia are characteristic of many flies.
Often, the larva of the last instar weaves a cocoon before pupation. So, for example, a caterpillar of silkworm butterflies secretes silk from the silk glands, from which it spins a dense cocoon. Inside such a cocoon is a covered chrysalis. And in some Hymenoptera - ants, as well as in the Neuroptera, there is an open, or free, pupa inside the cocoon. In larvae of lacewings, such as golden eyes, threads for the cocoon are produced by malpighian vessels and excreted from the anus.
Physiology of metamorphosis. In the process of metamorphosis, two interrelated processes occur: histolysis and histogenesis. Histolysis is the breakdown of tissues of larval organs, and histogenesis is the formation of organs of an adult insect. In insects with incomplete metamorphosis, these processes proceed gradually in the nymph phase, and in insects with complete metamorphosis, in the pupal phase.
Histolysis occurs due to the activity of phagocytes and enzymes. In this case, first of all, the fat body, larval muscles and some other organs are destroyed, which are converted into a nutrient substrate consumed by developing tissues.
Histogenesis, or the formation of the organs of an adult insect, occurs mainly due to the development of imaginal discs - rudiments from undifferentiated cells. The imaginal discs are laid down during the larval phase and even during embryogenesis and represent internal rudiments. Imaginal discs develop eyes, wings, mouth apparatus, legs, as well as internal organs: muscles, gonads. The digestive system, Malpighian vessels, and tracheae are not destroyed, but are strongly differentiated in the process of metamorphosis. The least metamorphosed heart and nervous system. However, during metamorphosis in the nervous system, the process of oligomerization (fusion) of ganglia is often observed.
The process of metamorphosis is controlled by the endocrine glands (Fig. 329). neurosecretory cells brain secrete hormones that stimulate activity cardiac bodies, whose hormones through the hemolymph stimulate prothoracic(prothoracic) glands that secrete molting hormone - ecdysone. Ecdysone promotes the molting process: partial dissolution and exfoliation of the old cuticle, as well as the formation of a new one.
In the process of metamorphosis, the activity adjacent bodies producing juvenile hormone. At high concentrations, the molting of the larva leads to the formation of a larva
next age. As the larvae grow, the activity of the adjacent bodies weakens and the concentration of juvenile hormone falls, and the prothoracic glands gradually degenerate. This leads to the fact that the larvae molt into the pupal phase, and then the adult.
The artificial transplantation of adjacent bodies, for example, into a locust nymph of the last age, contributes to the fact that it does not molt into the adult phase, but into a larger larva of additional age. In the adult phase, the juvenile hormone controls the development of the gonads, and the hormone ecdysone is no longer produced due to the reduction of the prothoracic glands.
Origin of metamorphosis. There are several hypotheses about the origin of metamorphosis in insects. For a long time there were disputes about which insects are more evolutionarily advanced - with complete or incomplete transformation. On the one hand, insect nymphs with incomplete metamorphosis are more progressively developed than insect larvae with complete metamorphosis, on the other hand, the latter have an advanced pupal phase.
At present, this contradiction has been removed by the hypothesis of the origin of metamorphosis by G. S. Gilyarov, A. A. Zakhvatkin and A. G. Sharov. According to this hypothesis, both forms of metamorphosis in insects developed independently of a simpler type of development - protomorphosis, observed in primary wingless insects, for example, in bristle-tailed insects (Thysanura).
With protomorphosis, development is direct, with many molts observed in the larval phase, and then in the imaginal state. All phases of development of these insects occur in the same environment.
It is assumed that in the process of evolution, insects passed from a semi-hidden existence in the upper soil layer to living on its surface and on plants. This transition to a new habitat was crowned with a major aromorphosis - the development of wings and flight.
The development of open habitats was reflected in the evolution of the individual development of insects. The evolution of insect ontogeny apparently followed two main directions.
In one case, the process of embryonization of development was going on, leading to the hatching of insects from eggs rich in yolk in the later phases of development. This led to the imaginization of larvae with the formation of nymphs. This is how insects with incomplete metamorphosis developed. This path of evolution led to the progressive development of larvae leading a similar way of life with adults.
In the other case, on the contrary, a process of deembryonization of development took place, i.e., eggs poor in yolk were hatched at earlier stages of development. This led to the morphoecological divergence of insect larvae and adults. The larvae have simplified and adapted
to live in a more protected environment, performing the function of nutrition, and adults began to mainly perform the function of reproduction and settlement. In addition to the deembryonization of the development of insect larvae with complete transformation, they have developed many provisional adaptations to various conditions of existence. Thus, amphigenesis (divergence) occurred in the evolution of larvae and adults in insects with complete metamorphosis. Amphigenesis of larvae and adults turned out to be very deep in terms of morphological adaptations, which created serious contradictions in ontogeny. They were successfully resolved by the emergence of the pupal phase, during which a radical restructuring of the larval organization to the imaginal takes place. This allowed insects with complete transformation to master a wider range of ecological niches and achieve an unprecedented flourishing among animals on Earth.
Insect breeding. For most insects, bisexual sexual reproduction is characteristic. Many species show sexual dimorphism. For example, male stag beetles have mandibles modified into horns, and male rhinoceros beetles have a horn on their heads and humps on their pronotum. This is due to the mating behavior of these species, accompanied by the struggle of males for a female. The relationship between the sexes in different species is extremely diverse. Male Diptera of the Dolichopodidae family bring a "gift" to the female - a caught fly and perform a dance with leg mirrors. Female praying mantises are predatory and eat the male during mating.
Most insects lay eggs, but live births are often observed. In this case, the eggs develop in the female genital tract and she gives birth to larvae. For example, blowflies-sarcophagi (Sarcophagidae) lay live larvae on the meat, the development of which proceeds very quickly. No wonder in ancient times it was believed that worms in meat spontaneously generated. It is possible not to notice how a blowfly visited openly lying meat, and suddenly discover white larvae that suddenly appeared.
The viviparous species also include the sheep bloodsucker fly, some beetles that live in caves.
In addition to bisexual sexual reproduction, a number of insects exhibit parthenogenesis - development without fertilization. There are many species from different orders of insects, which are characterized by parthenogenesis. Parthenogenesis can be obligate - mandatory, then all individuals of the species are only females. So, in alpine conditions, in the north and in other unfavorable conditions, parthenogenetic beetles, orthoptera, earwigs, and lacewings are found. Parthenogenesis also occurs in bisexual species, when part of the eggs are laid fertilized, and part without fertilization. For example, bee drones develop from unfertilized eggs.
Similar parthenogenesis is also found in other Hymenoptera (ants, sawflies), termites, some bugs and beetles. And in aphids, for example, there is a change of generations in the life cycle: bisexual and parthenogenetic. In some cases, parthenogenesis can be optional (temporary), manifesting itself only under adverse conditions. Parthenogenesis in insects contributes to the maintenance of high populations.
A variant of parthenogenesis is pedogenesis - reproduction without fertilization in the larval phase of development. This is a special way of development of insects, when the maturation of the gonads is ahead of other organs. For example, some species of gall midges reproduce in the larval phase. Older larvae give birth to younger larvae. Pedogenesis was noted for one of the species of beetles, the larvae of which partially lay eggs and partially give birth to larvae. Pedogenesis, in addition to bisexual reproduction, increases the abundance of the species.
Life cycles of insects. Unlike ontogenesis, or the individual development of insects, the life cycle is the development of a species, which, as a rule, includes several types of ontogenies. Ontogeny is limited to the life of one individual from the egg to the onset of puberty and then natural death. The life cycle is a recurring part of the continuous development of a species. Thus, in the most typical case in insects, the life cycle consists of two conjugated and morphophysiologically different ontogenies of males and females, which reproduce sexually and reproduce their own kind. And in parthenogenetic species, the life cycle is characterized by only one type of female ontogeny.
The life cycles of insects are diverse in terms of types of reproduction, the composition of generations and their alternation. The following types of insect life cycles can be distinguished.
1. Life cycles without alternation of generations with bisexual sexual reproduction. This is the most common type of life cycle, characteristic of dimorphic species, consisting only of males and sexually reproducing females. These are the cycles of most beetles, butterflies, bedbugs.
2. Life cycles without alternation of generations with parthenogenetic reproduction. Such species are monomorphic, consisting only of parthenogenetic females that lay eggs without fertilization.
Parthenogenetic species are especially common among aphids, psyllids, and other homoptera. Parthenogenetic species of beetles, bugs, grasshoppers, and coccids are common in high-mountain conditions.
3. The rarest type of life cycle in insects is a cycle without alternation of generations with sexual reproduction of hermaphrodite species.
An American species of fly is known, consisting only of hermaphroditic individuals. In the early phases of development, the adults function as males, and in the later stages, as females. Therefore, all individuals lay eggs, which increases the abundance of the species.
4. Life cycles without alternation of generation with sexual reproduction and facultative parthenogenesis in polymorphic species, for example, in social insects. The species consists of sexual individuals - males and females, and fertile - working individuals that do not participate in reproduction. These species include bees, ants, and termites. Such life cycles are complicated by the fact that females lay, along with fertilized eggs, also parthenogenetic ones, from which, for example, bees develop haploid males - drones, and from fertilized ones - females and female workers. Some riders, thrips, coccids develop similarly.
In other species, facultative parthenogenesis manifests itself in a different way: not males, but females develop from unfertilized eggs. But in this case, the diploid set of chromosomes is restored in females by fusion of haploid nuclei. Such a development is known in some stick insects, locusts, sawflies, coccids.
5. Life cycles with alternation of sexual generation and parthenogenetic (heterogony). In many aphids and phylloxera, in addition to the sexual generation of winged males and females, there are several alternating generations of parthenogenetic females, winged or wingless.
6. Life cycles with alternation of sexual generation and several generations with pedogenesis. For example, in some gall midges, after sexual reproduction, in which males and females participate, parthenogenetic reproduction of larvae (pedogenesis) occurs. After several generations of breeding larvae, which die off after the live birth of their own kind, the last generation of larvae pupates and gives rise to winged females and males.
7. Life cycles with alternation of sexual generation (males and females) with asexual. After sexual reproduction, females lay fertilized eggs that undergo polyembryony. This is asexual reproduction in the embryonic phase. The egg undergoes crushing, and the embryo in the morula phase begins to multiply by budding. From one egg, several dozen embryos can form. Such
Thus, the classification of life cycles can be represented as follows.
I. No alternation of generations:
- 1) with bisexual sexual reproduction (May beetle);
- 2) with parthenogenetic reproduction (alpine beetles, grasshoppers);
- 3) with sexual reproduction of hermaphroditic individuals (American fly);
- 4) with sexual reproduction and partial parthenogenesis in polymorphic species (bees).
II. With alternation of generations:
- 1) heterogony: alternation of the sexual generation and several parthenogenetic (aphids, phylloxera);
- 2) heterogony: alternation of the sexual generation and several paedogenetic generations (some gall midges);
- 3) metagenesis: alternation of sexual generation with polyembryony (riders).
Seasonal cycles of insects. If the life cycle is understood as a cyclically repeating part of the morphogenesis of a species from one phase of development to the same phase, then the seasonal development cycle is understood as a characteristic of the development of a species during the seasons of one year (from winter to winter).
So, for example, the life cycle of the May beetle lasts 4-5 years (from egg to mature adults), and the seasonal cycle of this species is characterized by the fact that overwintered larvae pupate in spring and young beetles reproduce. In summer, autumn and winter, their larvae of different ages are found. The number of generations developing during the year is called voltness.
There are species that give several generations per year. These are multivoltaic species. For example, a housefly can give 2-3 generations per season and hibernates in the adult phase. Most insects are monovoltine, producing one generation per year.
Seasonal cycles of insects in nature are characterized by calendar periods of occurrence of various phases of development. Important features of the seasonal cycles of species are the timing of their active life and diapause (temporary delay in development) in winter or summer. The regulation of the life cycles of species in accordance with local seasonal phenomena is provided by environmental factors and the neurohumoral system of the body.
The embryonic, or embryonic, development of the worker bee, uterus, drone includes all the changes that occur under the egg shell, as a result of which a multicellular creature, the larva, is formed from a single-celled egg (Fig. 32). The egg just laid by the uterus, as a rule, is glued to the bottom of the cell with one end and stands vertically on it. The length of the egg is 1.6-1.8 mm, the width is 0.31-0.33 mm. It is slightly curved, of an elongated cylindrical shape, the end opposite from the place of attachment is slightly expanded. On
- On the 3rd day, the egg assumes an inclined position, and on the 3rd day, it lies on the bottom of the cell. At the free expanded end of the egg there is a tiny hole through which sperm enter from the uterine receptacle during the passage of the egg through the oviduct. This hole is called a micropyle. At other times, the micropyle is impermeable to both bacteria and viruses. The egg has anterior and posterior ends. The head of the larva develops in the free anterior end of the egg.
Embryonic development begins with the fragmentation of the nucleus. The daughter nuclei formed as a result of division are first randomly located inside the yolk, and then move with small areas of the cytoplasm to the periphery of the egg and form a continuous layer of cells - the blastoderm, which lines the entire egg shell from the inside. On the convex side of the egg, blastoderm cells begin to grow and multiply faster than the cells on the concave side. As a result, an embryonic streak is formed in the form of a wide strand. This is the beginning of the formation of the embryo. The germinal streak grows due to cell division and differentiation, tissue formation. As a result of these processes, the development of the larva occurs. In the second half of the 2nd day, the process of segmentation of the embryo begins. At the anterior end of the germinal strip, a preoral lobe (acron) is formed, on which a tubercle is formed - the rudiment of the upper lip. Below this tubercle, a recess appears - the future mouth. Then - the rudiments of the antennae. The segments of the thorax and abdomen detach from the head segment backwards. From the ventral (ventral) side of these segments, outgrowths appear - the rudiments of paired limbs. First it occurs on the chest, and later on the abdomen. At the same time, a distinct segmentation is found in the head section: the rudiments of five segments of the head are formed. In addition, breathing holes appear - stigmas, as well as the beginnings of the spinning glands and Malpighian vessels. As the segments of the body and limbs are formed, the separation of individual parts occurs.
Subsequently, the germinal streak increases in size. The reproduction of blastoderm cells leads to the invagination of the middle part of the germinal strip (the process of gastrulation), which descends deeper into the egg and separates from the blastoderm. In the process of gastrulation, first a longitudinal groove is formed on the surface of the germinal strip, and then the outer edges of the groove begin to grow towards each other. As a result, the future second germinal layer invaginates inside the germinal strip. When the edges of the groove close, the outer layer (ectoderm) is formed. Beneath it lies the inner layer (mesoderm). At the ends of the mesodermal layer, invaginations appear, consisting of groups of cells that form the third germinal layer (endoderm). The rudiments of individual organs begin to appear. Their development and differentiation continues until the larva hatches from the cell.
From the ectoderm, the body wall, the anterior and posterior intestines, the tracheal and nervous systems, and many glands are formed; from the endoderm - the rudiments of the sting, wings and limbs, the end sections of the genital tract, the nervous system and sensory organs; from the mesoderm - muscles, fat body, sex glands.
A fully formed embryo, which has essentially turned into a larva, fills the entire egg. She begins to move intensively, draw air into the trachea, swallow amniotic fluid, thereby increasing the volume of her body. The larva is released from the embryonic membranes, breaks the chorion (the secondary shell of the egg, formed by the transformation of the follicular epithelial cells) and hatches, that is, it comes out. Usually, the embryonic development of honey bees lasts about 3 days. Changes in temperature can increase or shorten the period of embryonic development.
After the release of the larva from the egg, postembryonic development begins, which is accompanied by metamorphosis (transformation).
Metamorphosis is a deep transformation in the structure of the organism, as a result of which the larva turns into an adult insect. Depending on the nature of postembryonic development in insects, two types of metamorphosis are distinguished:
incomplete (hemimetabolism), when the development of an insect is characterized by the passage of only three stages - eggs, larvae and the adult phase (adult);
complete (holometabolism), when the transition of the larva to the adult form is carried out at an intermediate stage - the pupal stage.
With incomplete transformation, the way of life of the early stages and adults is similar. The larva with this type of development is similar to an adult insect and, like it, has compound eyes, mouth organs, and then the rudiments of wings that are clearly visible from the outside. The rudiments of the wings increase in size with each subsequent molt, and in the last preimaginal age they may already cover several segments of the abdomen. In an insect of the last preimaginal age, the imaginal structures are fully developed, and an adult appears as a result of the last molt fully formed. Incomplete transformation is characteristic of orthoptera, bugs, homoptera, etc.
With complete metamorphosis, the entire development cycle is characterized by the passage of the stages of egg, larva, prepupa, pupa, and adult (Table 3).
- The duration of the stages of development of individuals of the bee colony, days
The transition from the larva to the adult takes place at the pupal stage. This is a non-nourishing stage during which tissues belonging to
ing earlier stages, imaginal structures are formed. In most insects, this stage is immobile. The pupa does not respond to external stimuli and is dressed in a thick covering. At the end of this stage, the shell bursts and an adult insect appears.
In insects, the regulation of metamorphosis is carried out by hormones produced by the endocrine glands (endocrine glands). Currently, three metamorphosis hormones are known that control postembryonic development: juvenile hormone, molting hormone (larval), and activation hormone. Juvenile hormone is produced and stored in the adjacent bodies, which are two small cell structures that lie behind the brain on the sides of the esophagus.
If the adjacent bodies are removed from a young larva, then pupation will follow the next molt, despite the fact that in its normal state it would have to do several more molts. This hormone determines the growth of larvae, controls the development of a number of organs and processes. For example, the fat body does not grow and function in the absence of this hormone.
The molting hormone, or ecdysone, is produced in the prothoracic glands, which are located in the prothorax of the larva above the first nerve ganglion near the first spiracle. It is synthesized in the body of the insect from. cholesterol. The hormone triggers the molting process and thus indirectly regulates growth and structure formation. The introduction of this hormone to insects causes the formation of swellings on giant chromosomes. The analysis of these swellings showed that an intensive process of RNA formation takes place in them. It is believed that the first result of the action of ecdysone is the activation of genes, then the stimulation of RNA biosynthesis and the formation of the corresponding enzymes that ensure the process of metamorphosis.
The activating hormone is produced in special neurosecretory cells, which are located in the anterior dorsal part of the supraesophageal ganglion. These glands are better developed in worker bees, less in queens, and even less in drones. The activation hormone affects the resumption of insect activity after each molt and stimulates the formation of two other hormones - ecdysone and juvenile.
The honey bee is an insect with complete metamorphosis (Fig. 33).
Larval stage. It is characterized by intensive nutrition and growth. The stage of the worker bee larva lasts 6 days, the queen - 5, the drone - 7. In the first days, the larvae of honey bees receive so much food that they swim on its surface. The basis of the feed is the secret of hypofarin-
| 1 g | 3 | 4 5 | 6 7 8 amp; | 10 | 11 12 | 13 14 15 16 17 18 19 20 | 21 |
| sh |
Day
Rice. 33. Stages of bee development
geal glands of nurse bees - a translucent white liquid. The milk secreted by the glands of nurse bees is highly nutritious. It contains carbohydrates, proteins, fats, minerals, vitamins of group B. Before giving food to the larva, the worker bee, lowering its head into the cell, repeatedly opens and closes the upper jaws. Then it touches the bottom of the cell with its jaws and releases liquid food. Other bees-nurse in the same way add the following portions of milk to the cell with the larva. For the entire larval stage, there are about 10 thousand visits to a cell with one larva.
The food of young larvae and older larvae is not the same. In the second period of life, the larva of the worker bee begins to receive gruel - a mixture that is prepared by nurse bees from honey and pollen.
During the larval stage, the linear dimensions of the worker bee larvae increase by more than 10 times. Thus, the length of a newly hatched larva is 1.6 mm, a 1-day old larva is 2.6 mm, a 2-day old larva is 6 mm, and by the end of the larval stage it is 17 mm (it almost completely occupies the bottom of the cell). The length of the uterus larva reaches 26.5 mm by the end of the stage.
Intensive nutrition provides the larvae with a significant increase in body weight. The hatched larva weighs about 0.1 mg. For the first
- days, the mass of a worker bee larva increases by 45 times, and the mass of a drone larva - by 85 times. By the end of the stage, the weight of the larvae of working individuals increases by 1565 times, and that of the mothers by 2926 times.
In appearance, the larva differs sharply from the adult bee (Fig. 34). It has a worm-like shape, soft covers of a whitish color. Its body consists of a head, a jointed trunk and an anal lobe.
The head of the larva is small, obtuse-conical, the base of the cone is formed by the head capsule, which is divided by a longitudinal suture.
Rice. 34. External (G) and internal (1G) structure of the bee larva "\u003e
Rice. 34. External (D) and internal (1D) structure of the bee larva:
A - general view; B - view from the side of the head; C - position of the adult larva at the bottom of the cell; G - side view; y - rudiments of antennae; vg - upper lip; vh - upper jaw; opzh - opening of the spinning gland; ng - lower lip; p - mouth; ndg - supraesophageal ganglion; PC - foregut; a - aorta; sk - middle intestine; zht - fat body; zpzh - the germ of the sex gland; c - heart; zk - hindgut; ms - malpighian vessel; bn - abdominal nerve chain; pzh - spinning iron; pdg - subesophageal ganglion
into two equal convex parts, the so-called cheeks. The apex of the cone at the front of the head is formed by the upper lip and other mouthparts. The part of the head, including the lower lip, the upper jaw, and the part of the head capsule to which they are attached, is separated from the head capsule by a deep groove that runs on each side between the base of the upper jaw and the maxilla and extends to the posterior borders of the head.
The mandibles extend from the head capsule in the form of cone-shaped organs. The antennal rudiments are located on the head capsule above the base of the mandibles in the form of round tubercles.
The larvae on the thoracic segments have no outer rudiments of either wings or legs. There are no compound eyes. However, under the larval cuticle, their rudiments are indicated, which are called imaginal buds. The imaginal buds of the legs are spindle-shaped and dissected by transverse grooves. The rudiments of the wings consist of two flat processes of the hypodermis bent inward. They are located on the sides of the second and third thoracic segments. The formation of such structures in an adult insect from imaginal buds allows the development of specialized structures in larvae, In the region of the eighth and ninth abdominal segments on the ventral side, the rudiments of the genital organs appear.
It should be noted that the development of wings, antennae, eyes, and legs does not occur through the restructuring of reduced larval structures; they develop from isolated areas of embryonic tissues that are in an inactive state, called rudiments.
The larvae differ from the adult bee in the structure of the internal organs (see Fig. 34). The central place in the body of the larva is occupied by the intestinal canal, which consists of three sections - the anterior, middle and hindgut. The foregut looks like a short thin tube, slightly expanded at the posterior end. The anterior intestine contains a small oral cavity, a short pharynx and esophagus. Honey goiter, characteristic of adult bees, is absent. In the wall of the foregut there are muscles that ensure the ingestion of liquid feed. At the junction of the esophagus and the midgut, there is a small annular fold that plays the role of a valve. It closes the lumen of the esophagus and prevents the contents of the midgut from flowing back into the esophagus. By the end of the larval stage, a thick septum forms in place of the valve, covering the midgut. 1
The midgut is the largest section of the intestine and occupies most of the body of the larva. A distinctive feature of this section is the absence of its connections with the hindgut, as a result of which undigested parts of the food remain in it throughout the entire larval stage. The connection of these two sections occurs only before spinning the cocoon, when the larva is already sealed. During this period, the membrane that separates the middle intestine from the hindgut breaks through and the feces pass into the hindgut, after which they are removed from the body to the bottom of the cell.
In the middle intestine, the larvae have Malpighian vessels. The honey bee has four of them. They are long, slightly convoluted tubes that run along the midgut. During the larval life, the Malpighian vessels are closed. Only by the 6th day of the larval stage, when they swell strongly from the decay products accumulated there, the posterior ends and contents break through, pass into the hindgut, and then are removed. Thus, the larva does not defecate during its growth and feeding.
The hindgut is small and is a narrow tube, curved at an acute angle and ending in the anus.
The heart of the larva consists of 12 chambers of the same structure (there are only five in an adult bee) and is a thin-walled tube located directly under the cuticle of the abdomen and chest. At the second segment of the thoracic region passes the aorta. It has the appearance of a tube, bent downwards, which then passes into the head and ends at the anterior surface of the brain, where the hemolymph pours into the body cavity. On the surface of the aorta is a dense network of tracheae.
Between all chambers of the heart there are openings through which the hemolymph enters the heart. Due to the fact that the thin front ends of each chamber protrude into the next chamber, no reverse flow of fluid is allowed.
The respiratory system of the larva is quite simple and is represented by two large longitudinal tracheal trunks running along the sides of the body, with small branches from them that spread throughout the body. On the sides of the segments there are 10 pairs of spiracles, which are hollows surrounded by narrow chitinous rings.
The nervous system consists of two simply arranged head ganglia (a large supraoesophageal and a small subglottal) and a ventral nerve cord extending along the underside of the chest and abdomen. The abdominal chain consists of 11 ganglia - 3 thoracic and 8 abdominal. The last abdominal ganglion is in the middle of the eleventh segment; it was formed by the fusion of the ganglia of the last three segments. The ganglia are connected to each other by paired connecti. The suboesophageal ganglion is also connected to the ventral nerve cord through two connectives.
The genital organs are present in the larva of the worker bee in its infancy and are represented by two ridges located in the eighth segment. Their length is on average 0.27 mm, and their width is 0.14 mm. During the entire larval stage, the development of ovarian primordia occurs. In the 2-day-old larva, they are insignificant in size; in the 6-day-old larva, they are already elongated and are located towards the caudal end through the ninth segment. By the end of the larval stage, the number of egg tubes in a worker bee reaches 130-150. During the subsequent stages, when the larva turns into a pupa, the larval tissues disintegrate and the number of egg tubes in the worker bee decreases from 20 to 3, while in the uterus, the ovaries continue to develop in the pupal stage.
The fat body is well developed in the larva, where the accumulation of nutrients occurs. In older larvae, the fat body reaches 60% of body weight and fills almost the entire cavity between the midgut and cuticle of the larva. The fat body is a concentration of fats, proteins and carbohydrates, due to which the pupa develops in the future. The fat body, in addition to the actual fat cells, also consists of excretory cells and enocytes that trap uric acid salts.
The spinning gland is present only in the larvae. The secretion of this gland serves as material for spinning the cocoon before the transition of the larva to the pupal stage. The spinning gland looks like two long tubes, which are placed under the middle intestine from the ventral side. In the anterior chag, both tubes are connected into an unpaired excretory duct, which opens under the oral opening at the tip of the lower lip.
During the entire stage, the larva develops rudiments of a stinger, one pair of which is located on the eleventh segment and two pairs on the twelfth. There is an rudiment of the genital opening between them by segments. The rudiment of a large poisonous gland appears only towards the end of the larval stage, and at the beginning of the pupal stage, a small poisonous gland forms from the deepening of the cover on the ninth segment.
On the 5-6th day after the release of the larvae from the egg, the bees begin to seal the cell with a wax cap. In addition to wax, the composition of the material that makes up the cap includes pollen, water and papery mass. The lid turns out to be porous, due to which the air necessary for the respiration of the larva, and then the pupa, penetrates into the cell. From this moment on, the larva completes its growth and stops feeding.
As soon as the cell is sealed, the larva straightens up in it, the intestinal walls contract and the undigested food residues, having first broken through the thin wall of the middle intestine, pass into the thick one, and from there outward. The larva deposits excrement in one of the corners of the cell. After that, the larva spins a cocoon, i.e., braids the inner surface of the cell. When spinning a cocoon, the larva uses the secretion of the spinning gland, Malpighian vessels and sticky secretions from the walls of its body. The spinning process was studied and described in detail by Velich (1930).
While spinning the cocoon, the larva makes quick rhythmic trembling movements; starting from the head, the wave moves through the segments. In 1 min, the larva makes up to 280 movements. Simultaneously with trembling, the larva makes rocking movements of the head with the anterior end of the body. They are in the nature of dashed, elliptical, irregular curves from top to bottom or from right to left. At the same time, a transparent substance flows out of the opening with a valve on the lower lip, which the larva secretes in the form of one thick or several thin threads. Within 2 days, the larva, turning, strengthens the walls of the cocoon. When spinning a cocoon, the behavior of the larvae of worker bees, drones, and queens was similar, the difference being only in the shape of the cocoon. It depends on the size of the cell in which the larva is located. For example, in a worker bee, the cocoon corresponds to the shape of a hexagonal prism, the bottom of which is a three-sided pyramid, and the lid is an exact imprint of a sealed bee cell. The cocoon after the exit of the young bee is not removed. Due to the fact that dozens of generations are displayed in one cell, the diameter of the cell narrows over time. So, if a normal cell of a freshly built comb has a volume of 0.282 cm3 and a bottom thickness of 0.22 mm, then after 20 generations of bee individuals, the cubature of the cell decreases to 0.248 cm3, and the bottom thickens to 1.44 mm.
The larva of the worker bee and the queen spins a cocoon for 2 days, and the larva of the drone for 3 days. She then molts a fifth time. This molt is preceded by a number of changes in the organism of the larva. The rudiments of all appendages located under the cuticle begin to turn inside out or move outward and turn into external parts of the body. These organs gradually take the form characteristic of the pupa. The head, mouthparts, and legs turn from anterior to posterior-abdominal. After spinning the cocoon, the larva straightens up and freezes. The preparatory process for the pupal stage begins, which requires considerable time. This period is called the prepupal stage.
Prepupal stage. The duration of the prepupa stage for the worker bee is 3 days, for the queen - 2, for the drone - 4 days. This period is characterized by significant transformations in the structure of the larva. The larval cuticle is completely separated from the pupal cuticle under the influence of a special molting hormone ecdysone. An exception is the tracheal connections between larval and pupal spiracles. The head is already 2/3 the size of the adult head. The eyes are enlarged, and their surface becomes folded and wrinkled. The chest is separated from the head by a clearly visible constriction. The wings begin to grow in breadth, at this stage they are wrinkled and tightly pressed to the chest. The mouthparts elongate and merge into a compact group.
The segments of the chest and abdomen are divided along a horizontal line into two halves - dorsal and abdominal, gradually acquiring the structure characteristic of an adult.
In addition, internal organs are subject to decay. This process is called histolysis. It is accompanied by the penetration and introduction into the tissues of blood cells - phagocytes. The sources of energy necessary for the chemical reactions underlying these processes are the reserves of fats, glycogen and other carbohydrates in the fatty body of the larva, the sugar of the hemolymph and muscles. During histolysis, hemocytes begin to function as devouring cells, i.e., phagocytes, and enzymes convert the fat body, most of the muscles of the larva, into a nutrient substrate delivered by hemolymph to growing tissues. In the later stages of histolysis, the phagocytes themselves are taken up by the new developing tissues. The muscular system undergoes the greatest changes, as a result of which the prepupa is immobilized.
A honey goiter with a valve appears in the anterior intestine, the middle intestine becomes loop-like and acquires a folded structure. In the hindgut, two sections appear - the small intestine and the rectum. The number of tubules of Malpighian vessels increases. Partial fusion of ganglia of the neural chain is observed, as a result of which the pupa has seven ganglia of the abdominal chain instead of 11 ganglia of the larva. The fat body is reduced in size.
The prepupa stage ends with the shedding of the old cuticle, and the pupa emerges from under it.
Pupa stage. The entire surface of the body of the newly emerged pupa is covered with folds, but not wrinkled. In its structure, the pupa is similar to an adult bee, although, unlike the latter, it is motionless, its body is devoid of pigmentation (white). However, internal life processes do not slow down, but proceed with great efficiency. The pupa immediately after the completion of the last molt is covered with a cuticle. The pupal stage is characterized by profound transformations. The source of plastic substances for the formation of new organs and tissues, or for histogenesis, are the decay products carried by hemocytes. The spinning glands, the digestive canal, the fat body, and the muscles completely disintegrate.
Instead of decayed organs and tissues of the larva, new organs of an adult insect are formed.
The head is much longer and expanded almost to the size of the adult head. There is a bridge between the expanded head and the narrowed prothorax. However, it is absent between the thoracic and abdominal segments. The pupa does not yet have wings, but the antennae, proboscis and legs are already clearly marked.
Some of the elements of the fat body during this period are scattered in the head and chest, and some are concentrated in the abdomen. With age, the elements of the fat body are more evenly distributed in the head, chest and abdomen. The structure of the stomach is changing.
The heart changes little: the number of chambers decreases from 12 to five. Throughout the pupal stage, the heart does not stop functioning, since the process of metamorphosis requires the movement of hemolymph. The mass of the supraesophageal ganglion in the head increases. This happens due to undifferentiated nerve cells. In addition, the nerve ganglia in the chest merge, as well as the last four ganglia at the end of the abdomen.
The rudiments of the gonads are not destroyed. Their growth at the initial stage is associated with the disintegration of the cells of the fat body and the release of plastic material in the form of albumides. As they develop, they come into contact with parts of the gonadal ducts arising from the ectoderm.
One of the indicators of the internal life of the chrysalis is its breathing.
The respiratory system undergoes a number of changes during histolysis and histogenesis. So, the first and third thoracic spiracles increase, and the second, on the contrary, decreases. Destroyed, trachea. New tracheae, as well as air sacs, are formed from imaginal rudiments, which are located on the tracheal trunks. 1
Compound eyes grow from buds on the sides of the head. As mentioned above, the pupa is initially white. During the development in the process of formation of various organs, the color of the outer integument of the pupa changes in the following sequence: a day after pupation, the compound eyes are still pure white, after 2 days a yellowish tint appears, on the 14th day they become pinkish in color, and by the 16th day - dark purple.
The joints of the legs are yellowish at first, from the 18th day the joints and claws on the legs acquire a yellow-brown color, the heels and mouthparts are somewhat lighter. The chest becomes ivory. From the 19th day, the chest begins to darken slightly. Belly ivory. The ends of the segments of the legs are brown. A more significant darkening is observed at the end of the chin and at the base of the upper jaw. The eyes turn purple. On the 20th day, the color of the entire pupa is dark gray.
Imago stage. At the end of the pupal stage, the pupal skin is shed, from under which a fully formed bee emerges. With its mandibles, the bee gnaws through the lid of the cell and emerges onto the surface of the comb. A newly hatched bee, compared to an older bee, has a softer chitinous cover and, in addition, its body is densely covered with hairs. Over time, bees lose part of their hairline, and chitin becomes much harder. During the imaginal period, no significant changes in the external structures of bee individuals occur. "
Metabolism in the postembryonic period. The larva and pupa undergo intensive metabolic processes, as a result of which the nutrients necessary for the formation of an adult bee accumulate.
According to Strauss (1911), a feature of the metabolism of bees is the accumulation of significant amounts of glycogen as energy for metamorphosis. By the end of the larval stage, the glycogen content reaches 30% of the dry mass of the larva. At the same time, there is an increase in the amount of fat, the growth of fat cells.
First of all, glucose is consumed in the body of bees. Its total content in the body of worker bee larvae is twice as high as in the body of mother larvae. When the glucose content in the hemolymph falls below the permissible value, complex carbohydrates begin to break down and be used. The highest concentration of glucose is observed in the middle of the open stage.
It should be noted that the activity of most of the studied enzymes is highest in young larvae. Then it decreases and reaches a minimum in the pupal stage. At the same time, the development of enzyme activity in worker bees is more significant than in queens. There is an opinion that during the period of postembryonic development, queens have a more aerobic carbohydrate metabolism compared to working individuals, and the accelerated development of queens is explained by the higher activity of a number of enzymes. Differences in the quantity and quality of mitochondria (cellular organelles) in uterine larvae older than
- day old compared with those of worker bee larvae of the same age. In accordance with this, some differences are also observed in some oxidative processes occurring in mitochondria.
In the queen larva, the oxygen consumption curve goes up until the cell is sealed. However, after sealing, the oxygen uptake curve drops (which coincides with cocoon spinning) and reaches a minimum in the initial pupal stage. The level of oxygen consumption per unit mass in queens is the lowest compared to worker bees and drones.
The amount of oxygen absorbed per individual in drones in the postembryonic period is higher than in queens and worker bees.
The respiratory coefficient, which characterizes the substrate used as an energy source during the entire stage of development of the larva in a worker bee, changes, and its value exceeds one. If the value of the respiratory coefficient is higher than one, then this indicates the intensity of the processes associated with the transition of oxygen-rich compounds to oxygen-poor compounds.
The increase in the respiratory coefficient in the second period of larval development coincides with the time of transition to a qualitatively different type of food, mainly carbohydrate. After the sealing of the cell and until the pupation of the larvae, the respiratory coefficient is set at 0.9.
The main sources of energy in the sealed larva and prepupa are carbohydrates deposited in the form of glycogen, and in the second half of the pupal stage, fats.
In the uterus, unlike the worker bee, the respiratory coefficient decreases by the 2nd day of larval development, which is characteristic of fat metabolism. On the 4-5th day of the same stage, the respiratory coefficient is almost equal to the value of the respiratory coefficient of the worker bee larvae, which indicates the transformation of carbohydrates into fat. In the sealed larval stage, the respiratory quotient is below one, indicating the breakdown of carbohydrates.
From the foregoing, we can conclude that gas exchange in honey bees is characterized by a number of patterns inherent in insects. During the development of larvae, the intensity of respiration, expressed as the amount of oxygen consumed per unit mass, decreases. However, the amount of oxygen consumed per larva increases, reaching a maximum before pupation. At the pupal stage, gas exchange decreases, increasing towards the exit of the adult. At the same time, along with similarities, there are differences in the respiratory exchange of the three forms of honey bees.
In insects, development is divided into two periods - embryonic and postembryonic.
Embryonic development. Insect egg cells are mostly oval in shape and very rich in yolk, which fills the entire central part of the egg. Accordingly, crushing of the egg is partial, superficial; as a result, a continuous layer of identical cells, the blastoderm, is formed, covering the central yolk. At the same time, part of the nuclei remains in the yolk mass: in the future, special yolk cells are formed at their expense. On the future ventral side of the embryo, the blastoderm thickens. This area is called the germinal streak; in the future, at its expense, the main part of the body of the embryo is formed.
In the region of the germinal strip, differentiation processes begin. As a result of reproduction and penetration into the depths of the blastoderm cells, two strips of cells are formed - mesodermal stripes, and the blastoderm itself becomes the ectoderm. Later, the mesodermal streaks divide into paired primordia, giving rise to a double row of coelomic sacs. In higher insects (neg. Diptera) segmentation of mesodermal stripes is suppressed and separation of independent coeloms does not occur.
The question of the formation of the endoderm in insects has not yet been fully resolved. In many forms, the vitelline cells appear to be the rudiment of the endoderm. At Diptera and some other insects, two groups of cells are separated at the anterior and posterior ends of the germinal strip, which are also considered as the beginnings of the endoderm.
Even at the very beginning of the laying of the germ layers, the process of formation of embryonic membranes, which is extremely characteristic of the development of insects, begins.
In the simplest and most widespread case, along the periphery of the germinal strip, the ectoderm first forms a roller, and then a two-layer fold, gradually growing from all sides to the germinal strip, which at the same time is somewhat immersed in the yolk. Finally, the edges of the fold close over the rudiment of the embryo, leaving between themselves and it the amniotic cavity occupied by fluid, separated from the external environment by a double membrane. The outer layer of the membrane occurs as a result of the fusion of the outer layers of the ectodermal fold closing over the embryo and is called the serosa, the inner layer from the inner layer of the same fold, the latter directly limits the amniotic cavity and is called the amnion. Serosa and amnion protect the embryo from damage.
Meanwhile, the germinal streak experiences segmentation, and the segments are isolated, as in other Arthropoda and rings, in order from front to back. In the head section, the ophthalmic and antennal lobes with the corresponding rudiments, the intercalary segment, which subsequently undergoes reduction, and 3 segments of the oral limbs, are laid. Further posteriorly, 3 thoracic, bearing the rudiments of limbs, and 11 abdominal segments are differentiated.
As for the differentiation of internal organs, in its main features it consists in the following. The foregut and hindgut are formed by two ectodermal invaginations. The question of the origin of the midgut is more complicated. Insects are characterized by an amazing variety of ways and sources of the formation of the latter. In some cases, it is formed due to yolk endodermal cells, in others, ectodermal elements take part in the formation of the midgut - special cells localized at the blindly closed ends of the rudiments of the anterior and posterior intestines. And, finally, in a number of species (parthenogenetic female aphids, some ichneumons), the midgut is formed only by the ectoderm due to the strong growth of the anlage of the foregut, which reaches the posterior end of the body. Endodermal elements degenerate at the same time. Malpighian vessels arise from the ectoderm of the hindgut. The nervous system is laid down in the form of a ventral ridge of ectoderm, which then sinks under the integument. Coelomic sacs disintegrate, forming a mixed body cavity - a mixocoel. The cellular elements released at the same time give rise to muscles, heart, fat body and some other organs. Primary germ cells are laid down very early, sometimes even during the period when the yolk is surrounded by a single layer of blastoderm.
The embryo grows, the nutritious yolk is constantly consumed, and the embryonic membranes burst, wrap on the dorsal side in the form of folds, which undergo degeneration. The embryo is ready to emerge from the egg, and with this the embryonic development of the insect ends.
Postembryonic development. In insects, a young animal or a larva with a full number of segments emerges from the egg. The only exceptions are primitive cryptomaxillaries, belonging to neg. Protura, in which juveniles differ from adults in the absence of three posterior abdominal segments. The latter are formed from the rear growth zone sequentially after the first molts. Thus, development without anamorphosis is typical for insects, and only in protur is residual anamorphosis found.
The postembryonic development of insects is characterized by considerable diversity, which is largely determined by the biological characteristics of representatives of this group of animals.
In the most primitive forms (subcl. Entognatha) the growth and development of a young organism is not accompanied by significant structural changes. Juveniles in all main features are similar to the mother's organism, i.e., the larval stage and metamorphosis are absent. In essence, in this case we are dealing with direct development. Winged insects, which are higher in evolutionary terms, on the contrary, have metamorphosis, and according to the nature of the transformation, they are divided into hemimetabolic - with incomplete transformation and holometabolic - with complete metamorphosis. Incomplete metamorphosis is characteristic of lower organized groups of winged insects: orthoptera, cockroaches, bugs, mayflies, dragonflies. With incomplete transformation, the larva hatching from the egg in general appearance is already in many respects similar to the adult organism, differing from it mainly in the embryonic state of the wings, the underdevelopment of secondary sexual characteristics, and often the presence of some provisional, purely larval organs: for example, tracheal gills in mayfly larvae, etc. Wings are at first only in the form of barely visible outer rudiments. The smoothing of the difference between the larva and the adult organism occurs gradually, accompanied by a number of molts.
With each subsequent molt, the larva becomes more and more similar to the adult stage of insects, or adults (imago), the rudiments of wings increase, etc.
In some primitive insects, the number of molts is very large and can reach 25-30 (mayflies). Representatives of other detachments have fewer lines (4-5).

In holometabolic insects, the development is different. The larva differs sharply from the adult animal: it does not have external rudiments of wings at all, it often has a different number of limbs, etc. The larval characters are preserved during a series of molts, i.e., during the entire larval period of life. The larvae of holometabolic insects are extremely diverse. The larva differs from the imago in the presence of a number of primitive features: a greater homonomy of dissection, the absence of wings, a weak development of the sense organs (there are no compound eyes, but only ocelli, antennas are very small, etc.), and in many cases also the device and number of limbs. Very often, the mouth limbs at the larval age are arranged according to one type (gnawing in butterfly caterpillars), in adults - differently (sucking mouthparts in adult butterflies).
As already mentioned, the structure characteristic of the larva is preserved throughout the entire larval period: the transformation does not occur gradually, but the changes necessary for metamorphosis are postponed to the time of the last larval molt. During this period, such fundamental transformations of the organism of the larva take place that the continuation of the usual physiological functions, especially nutrition and movement, becomes difficult, even impossible. This is reflected in the fact that the larva, after the last molt, goes into a state of rest - it becomes a chrysalis. The pupa constitutes the most characteristic stage of the holometabolic transformation. The more complete the state of rest, the better the complete transformation is expressed.
The individual development of an insect or ontogenesis can be divided into 2 periods - development inside the egg, or embryonic, and development after leaving the egg, or postembryonic.
Most insects lay eggs. An insect egg is a cell that contains a nucleus, protoplasm, deutoplasm, or yolk, necessary for the nutrition and development of the embryo. In addition, insect eggs often contain symbiotic microorganisms obtained from the mother through the ovaries (i.e., transovarially). The egg is covered on the outside with a chorion - a shell. The chorion sometimes has a distinct microsculpture, which serves as a reliable sign for distinguishing genera and even species of insects according to the phase of the egg. Under the chorion lies the true or yolk membrane of the egg. With the formation of the chorion, an opening is left to the micropyle, which serves for the passage of the spermatozoon during fertilization. The egg has anterior and posterior ends that correspond to the position of the embryo. In the ovary, the anterior pole of the egg is directed towards the mother's head, respectively, the dorsal and ventral sides of the egg are located.
The size and appearance of the eggs are varied. The size is 0.02 - 0.03 mm for aphids and thrips, 8-10 mm for locusts. The eggs are smooth in aphids, ribbed in scoops, whites, sometimes equipped with a lid, covered with wrinkles.
Egg shape. Oval - beetles, butterflies - elongated, hemispherical - scoops, bottle-shaped - diurnal butterflies, barrel-shaped - bugs, have a stem - lacewings, suckers. Oviposition: singly, in groups, openly or in substrate, may be protected, covered with gonadal secretions.
Embryonic development begins with the fragmentation of the nucleus and the movement of the resulting daughter nuclei with small areas of protoplasm to the periphery of the egg. Here, from the mass of daughter nuclei, a continuous layer of cells is formed - the blastoderm. Later, the blastoderm differentiates into the germinal and extra-embryonic zones: the cells of the latter do not participate in the formation of the embryo, while the cells of the germinal zone begin to divide more intensively and form an embryonic band on the ventral side of the egg, which then protrudes and ectoderm and mesoderm arise.
The development of the embryo is accompanied by blastokinesis: embryonic membranes are formed, the head segments, thoracic, and abdominal are separated. The foundations of the internal structure begin to be laid with the formation of the mesoderm. From the ectoderm come all the outer covers - then the mouth and anus, the anterior and posterior intestines. The mesoderm gives rise to the muscular system, fat body, dorsal vessel, sheath of the sex glands.
The embryo begins to make movements, captures air, liquid from the egg, the chorion breaks and the embryo, which has turned into a larva, comes out.
2. The reproduction of insects has a number of features that constitute an essential aspect of their biology and require consideration. These features include methods of reproduction, additional nutrition, meeting of the sexes and fertilization, sexual productivity.
Reproduction methods .
Reproduction in most insects is accompanied by mating and fertilization, i.e. associated with the participation of two sexes - therefore it is called bisexual (bisexual) gamogenetic. Most insects lay eggs, but there are other methods of reproduction: live birth, parthenogenesis, pedogenesis, polyembryony. Live birth is that embryonic development is completed in the body of the mother, therefore, instead of laying eggs, larvae (aphids, gadfly, bloodsucking flies) are born into the world. The larvae hatch in the egg tubes or while passing through the oviducts.
Parthenogenesis - virgin reproduction is characterized by the absence of fertilization and is observed in both oviparous and viviparous insects.
Forms of parthenogenesis:
1. arrhenotokia- only males develop from unfertilized eggs; 2) thelytokia - only females develop from unfertilized eggs; 3) amphitoky- males and females develop from unfertilized eggs.
Parthenogenesis can be facultative, permanent and cyclic.
Facultative parthenogenesis or spadic manifests itself inconsistently, occurs under external influences or at a certain physiological state of the egg-laying female (gypsy moth, poplar hawk moth).
Constant parthenogenesis of the arrhenotoky type is typical of social insects. In bees, unfertilized eggs always develop into males, and from fertilized eggs, females. Permanent parthenogenesis of the thelytoky type is observed in insects that do not have a colonial way of life. So, in many worms, thrips, sawflies, and some riders, males are absent, reproduction always occurs through parthenogenesis, and only females hatch from unfertilized eggs.
Parthenogenesis plays a very important role in the life of insects. Thanks to parthenogenesis and its modifications, the reproductive potential of the species increases dramatically: with parthenogenetic reproduction, the chances of the species to survive under adverse conditions and during settlement are significantly increased.
3. After leaving the egg, the postembryonic development of the insect begins. This period is not a simple growth and increase in body size, but is characterized by the transition of the body from one phase to another. This type of individual development is called metamorphosis or development with transformation.
The essence of metamorphosis lies in the fact that a developing individual undergoes a significant restructuring of its morphological organization and features of biology during its life. In this regard, there is a differentiation of postembryonic development into 2 phases - larval and adult. In the larval phase, the growth and development of individuals occurs, in the imago phase, reproduction and settling. In other cases, an intermediate phase occurs between these phases - the pupa. There are 2 main types of metamorphosis - incomplete and complete transformation. incomplete conversion or hemimetamorphosis 3 phases - egg, larva, adult. The larvae are similar to adults (compound eyes, mouthparts, rudiments of wings, a similar way of life), imago-like. With complete (holometamorphosis) 4 phases. The larvae do not look like adults at all, there are no complex eyes, wings, they live in different conditions.
Some have ventral legs, a different mouth apparatus, others have silk or spider glands.
Modifications of incomplete transformation: a) hypomorphosis - a simplified incomplete transformation and is characteristic of winged insects that have lost wings (secondary wingless lice, lice, wingless representatives of locusts, grasshoppers, crickets, cockroaches, stick insects, hay eaters, bedbugs); b) hypermorphosis - a complicated incomplete cycle (aleurodids, thrips, coccid males). The appearance at the end of the phase of the larvae of the resting state.
Anamorphosis- larvae are similar to adults but have a smaller number of abdominal segments; as the larva develops, additional segments grow at the top of the abdomen, but the full number reaches only in the adult phase.
protomorphosis– molting in the adult state, some similarity of the larva to the adult phase, but the absence of division of the larva into the thorax and abdomen (podura, two-tailed, bristle-tailed).
hypermetamorphosis- complication of complete transformation (the presence of 2 forms of larvae, and sometimes pupae (blisters)).
4. The larval phase is immediately after hatching from the egg. Initially light, after hatching the larva is colorless, has soft covers. In openly living insects, the larva quickly darkens and hardens. Feeding stimuli are created after the digestion of the remains of the embryonic yolk and the excretion of excrement. The larva enters into a pair of enhanced nutrition, growth and development. Their growth and development are accompanied by periodic molts, shedding of the skin cuticle - due to molts, the body increases and major changes occur. The number of links varies 3 (flies), 4-5 (bugs), 25-30 (padenki).
After each molt, the larva enters the next stage, or age, therefore, the molts share the age of the larvae. In accordance with the number of molts, the number of larval instars is also found. Determining the age of molts (necessary for planning the timing of the fight) is carried out according to the following features: antennae in locusts, in an increase in body size (size of the width of the head capsule) in butterfly caterpillars and beetle larvae. Larvae are of 2 types - adult (primary) and secondary.
Naiads - larvae with gills, live in the water (dragonflies, padyonki, stoneflies).
Campodeoid - mobile, dark-colored with dense integument and with 3 pairs of pectoral legs, a well-isolated prognathic head (ground beetles, swimmers, lacewings).
Worm-like - sedentary, light-colored, devoid of abdominal legs.
Caterpillars have a well-separated head skin, 3 pairs of thoracic legs, 2-8 pairs of abdominal ones.
pupal phase. This phase of development of metamorphosis is characteristic only of insects with a complete cycle. A feature of the pupa is the inability to feed and very often stay immobile. It lives off the reserves accumulated by the larva, and is often regarded as a dormant phase.
Outwardly, the pupa does not look like an adult, but it has a number of signs of the adult phase: external rudiments of wings, legs, antennae, compound eyes, etc. Histolysis and histogenesis occur in the pupal phase.
Histolysis- the disintegration of the internal organs of the larva, which is accompanied by the penetration and penetration into the tissues of blood cells - hemocytes. Hemocytes function as cell eaters, i.e. phagocytes, the increased activity of which leads to the destruction and absorption of tissue matter. Histolysis captures the muscular system, so the prepupa becomes immobile, affects the digestive system, but does not affect the nervous and reproductive systems, as well as the dorsal vessel.
Histogenesis- the process of creating tissues and organs of imaginal life. The source for the formation of these new tissues and organs are the products of histolysis. Histogenesis embraces the muscular and digestive systems, rearranging them for new, imaginal functions. The muscular system is rebuilt for new forms of movement for flight, the digestive system for new types of food.
During histogenesis, the main role is played by imaginal rudiments - groups of hypodermal cells from which certain tissues and organs arise.
The molting hormone is exidone, which is necessary for the normal development of larvae.
There are the following pupae types:
open pupae- free, only pressed to the body imaginal appendages (antennae, legs, wings), characteristic of beetles.
Pupae with movable mandibles - use movable upper jaws to exit the cocoon, and can themselves move (reticulate, caddisflies, toothed moths). Pupae with immobile mandibles are unable to use the latter when emerging from the cocoon (beetles, Hymenoptera, Fanoptera, many Diptera).
Hidden dolls covered with hardened, not shedding larval skin, which plays the role of a shell or false cocoon - puparia (flies).
covered- have imaginal appendages closely pressed and soldered to the body due to the fact that during the last molt the larva secretes a secret that, when hardened, covers the pupa with a hard shell (butterflies, ladybugs).
Before pupation, some larvae surround themselves with a cocoon (silk, cobweb). Sometimes the place of pupation is plant stems, earthen cradle, open pupation.
Imago phase.
The insects emerging from the pupa have signs of an adult, but at first the wings remain folded. Then they spread their wings, the integument thickens and acquires the color of a fully formed adult insect. In the adult phase, insects do not molt and are not capable of growth. The exceptions are padyonki, fools. The biological function of the adult phase is to disperse and reproduce. This is already a function of species life and is aimed at maintaining the existence of the species. Thanks to the wings, the ability of adult insects to settle greatly increases, and reproduction allows you to leave descendants in a new place.
The dispersal of adult insects occurs both by active and passive flight. Active flights are usually characteristic of large insects and are observed in a number of species of dragonflies, locusts, butterflies, beetles and are widespread. Passive flights are characteristic of aphids and flies.
The transition to adults is accompanied by external changes in body color, an increase in the size of the abdomen in the female, due to the development of ovaries filled with eggs - in fertilized females of termites and ants - dropping wings, immature locust - pink, mature - bright yellow.
Sexual dimorphism differences between males and females in a number of external, secondary sexual characteristics - in the shape and size of the antennae, body size, various structural details (rhinoceros beetle, deer beetle). Males are more mobile, lead a more open lifestyle.
Polymorphism - the existence of insects, outwardly different forms of the same species - (ants, bees, termites, males, females, workers, soldiers). Sexual polymorphism is controlled within the family and does not depend on the influence of external factors.
Ecological polymorphism arises under the influence of the external environment (long-winged, short-winged, wingless bugs).