A rovarok embrionális fejlődése jellemzően a tojás felületes összezúzásával kezdődik, ami az utolsó tápláló sárgája gazdag tartalmához kapcsolódik. A tojás felszínén képződött bastoderma a felület egy részén vastagabb; ez a megvastagodás csíracsíkot eredményez, és meghatározza az embrió ventrális oldalát.
A csíracsík széle mentén kialakult gyűrűs redő, amely a csíraszalag fölött együtt nő és együtt nő, két csírahártyaréteget ad - a belsőt (amnion) és a külsőt (serosa). A csíracsík sejtrétegéből a sárgája felőli oldalon,az elsődleges endoderma elkülönül - az endoderma és a mezoderma közös csírája. Ezt követően a csíracsík szegmentálódik.
A rovarok posztembrionális fejlődése igen változatos a rovarfiatalok létezésének sokfélesége és a hozzájuk való alkalmazkodás mértéke miatt. A posztembrionális fejlődésnek két fő típusa különböztethető meg: a nem teljes átalakulás, a hemimetabolizmus és a teljes átalakulás, a holometabolizmus. A holometabolizmust a fiatal egyedek mélyreható elváltozásai jellemzik, aminek következtében a fiatal egyedek szerkezetében élesen eltérnek a kifejlett rovartól (kifejlett). Ezért a fiatalok állapotából, azaz a lárva fázisból az imaginális fázisba való átmenethez a szervek radikális átstrukturálása szükséges; ezt a lárva és a képzelet fázisa közötti köztes fázis részvételével érik el, amely alatt a fiatal egyedek nyugalmi állapotban maradnak; ez a köztes fázis bábos. A bábállapot során a rovar belső szervei vagy többé-kevésbé átalakulnak, vagy teljes pusztuláson (histolisis) mennek keresztül, és újjáépülnek; ez utóbbi esetben a szerv újbóli kialakulása az embrionális természetű sejtekből (hisztoblasztokból) álló, úgynevezett imaginális korongok miatt következik be. Így a bábkorszak az aktív hisztolízis és hisztogenezis (szöveti neoplazmák) időszaka. Kívülről a báb az imaginális fázist képviseli jellegzetes szerveivel, amelyek azonban fejletlenek maradtak.
A magasabb rendű rovarok fejlődése holometabolikus a bogarakban, lepkékben, szúnyogokban, bolhákban, hártyafélékben és kétszárnyúakban.
A tökéletlen átalakulás - hemimetabolizmus - esetében nincs ilyen mély különbség a fiatal és a felnőtt fázis felépítésében: a juvenilis most jobban, majd valamivel kevésbé hasonlít szerkezetében az imaginális fázisra, végül azokban az esetekben, amikor a fiatal korból való átmenet fázis a felnőttre csak a régi burkolat (exuvia) növekedésére és leválására (a növekedéssel összefüggésben) redukálódik, amebolizmusról, vagyis az átalakulás hiányáról beszélnek; az amebolizmus példája a Collembola kialakulása az Apterygota körében. Általában azonban az egymást követő lárváknál, amelyeket vedlések választanak el egymástól, vagyis az előző kor burkolatának leszakadásával, a szervek alakjában is előfordulnak változások: például az antennák és a ceruszok hossza. relatíve nő, a szegmensek száma nő; egyes szervek újra megjelenhetnek, például a külső nemi szervek függelékei. Ilyen a fejlődés néhány apterygota esetében, mint például a Thysanura és a Diplura, valamint a legtöbb félmetabolikus Pterygota esetében. Egyes hemimetabolizmussal fejlődő Pterygota-ban (szitakötők, majálisok; kőlegyek és ízeltlábúak részei) a fejlődés bonyolult, mivel a fiatal egyedek más körülmények között élnek, mint az imágók; így a szitakötők, majálisok, kőlegyek fiatal egyedei a víztestekben fejlődnek, és ezzel összefüggésben vízlégzési szerveket - kopoltyúkat - szereznek, amelyek a lárva (ideiglenes) szervek példája. A létfeltételek a lárva más, a környezethez alkalmazkodó szerveinek fejlődésében is megmutatkoznak, például a fiatal kabóca mellső lábainak földalatti életével összefüggésben ásószervvé alakulása.
A rovarok fejlődése
A rovarok egyedfejlődése (ontogén) a tojásfázisban végbemenő embrionális fejlődésből és a posztembrionális fejlődésből - miután a lárva elhagyja a tojást a kifejlett fázis eléréséig - az imágóból áll.
Embrionális fejlődés. A rovartojások alakja változó a fejlődésük környezetéhez való alkalmazkodás miatt. Például a bogaraknál a tojások túlnyomórészt oválisak, és gyakrabban fejlődnek zárt szubsztrátumban; poloskákban - hordó alakú, az aljzathoz rögzítve; pillangókban - torony alakú vagy palack alakú; fűzősben (aranyszemű) tojások szárral. A tojásokat gyakran csoportosan rakják. A tojáskapcsolók nyitottak vagy zártak lehetnek. A nyílt tojásrakás példája a Colorado burgonyabogár tojásai, amelyeket a nőstény ragaszt a burgonyalevél alsó oldalára. A zárt tengelykapcsolók közé tartoznak az akridoidok tojáshüvelyei, amelyek a nőstények mellékmirigyeinek váladékai által cementált talajrészecskékből képződnek. A csótányok a női nemi traktusban képződő ootheca - tojáskapszulákba rakják le tojásaikat.
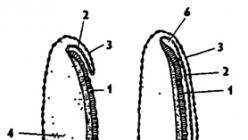
A rovartojásokat kívülről héj - chorion borítja, amely megvédi őket a kiszáradástól (337. ábra). A héj felületén van egy mikropila - egy kis lyuk, egy összetett "dugóval", amelynek belsejében egy tubulus található a spermiumok behatolásához a megtermékenyítés során. A chorion alatt vékony sárgája membrán található, alatta pedig sűrű citoplazmaréteg. A citoplazma központi része sárgával van kitöltve. A citoplazma tartalmazza a sejtmagot és a poláris testeket.
A zúzódás felületes. Kezdetben a sejtmag sokszor osztódik, a leánymagok a citoplazma szakaszaival a tojás perifériájára vándorolnak, membránnal borítják őket, és kialakul a sejt felszíni rétege - a blastoderma, a sárgája pedig a tojás közepén marad. tojás. A blastoderma ventrális felületén a sejtek magasabbak, és megvastagodást - a csíracsíkot - képeznek. A rovar embrionális fejlődésének ez a szakasza a blastulának felel meg.
A csíracsík sejtosztódása az embrió fejlődéséhez vezet. A csíracsík fokozatosan süllyed, kialakítva a hasi barázdát.
Rizs. 337. A rovartojás szerkezete (Bei-Bienko-ból): 1 - mikropila, 2 - chorion, 3 - sárgája membrán, 4 - mag, 5 - poláris testek, 6 - sárgája

A blastoderma barázda feletti redői összezáródnak, csírahártyák keletkeznek: serosa és amnion (338. ábra). Itt a magasabb gerincesekkel való konvergencia nyilvánul meg, amelyek szintén hasonló héjjal rendelkeznek. A kialakult magzatüreg miatt az embrió a tojás belsejében lebegő állapotban van, ami megbízhatóan megvédi a mechanikai sérülésektől, emellett a magzatüreget kitöltő folyadék elősegíti az embrió anyagcsere folyamatait.
A csíracsík további két rétegre különbözik: az alsó - ektoderma és a felső - endomezoderma. A különböző rovarfajok entomezodermája különböző módon alakulhat ki: invagináció vagy sejtbevándorlás útján.
A fejlődés következő szakaszában a csík ektodermális rétege oldalt felfelé hajlik, majd hátul záródik, és az embrió zárt falát alkotja. Amikor a test falai a háton összezáródnak, a tojássárgája és a tojássárgája sejtek egy része belép az embrió testébe. Az embrió teste falainak kialakulásával egyidejűleg két sejtcsoport válik el az entomesodermában a test elülső és hátsó végén. Ez a középbél két kezdete. Ezt követően ebből a két primordiából kezd kialakulni a középbél elülső és hátsó része, amelyek azután egyesülnek. Ugyanakkor az embrió testének elülső és hátsó végén az ektoderma mély invaginációi képződnek, amelyekből a bél elülső és hátsó része alakul ki. Ezután mindhárom szakasz összekapcsolódik, átmenő bélcsövet képezve.
A mezodermális csík cölomikus zsákok páros metamer rudimentumaira bomlik fel. Később azonban szétesnek, és a mezodermából kialakulnak az embrió izmai, a cölomikus hám szomatikus lapja, a szív, a zsírtest és az ivarmirigyek. A cölomikus epitélium zsigeri rétege a rovarokban nem képződik, és a testüreg keveredik - a mixocoel. A cölomikus rudimentumok egyesülnek az elsődleges testüreggel.
Később az ektodermából kialakul az idegrendszer és a légcsőrendszer. A malpighi erek a hátsó bél falaiból alakulnak ki.
A fejlődés során a rovar embriója szegmentálódik, amely először a test elülső részén, majd a hátsó részében nyilvánul meg. A fejrészben egy makkot szemmel, ajak- és

antennalebenyek, egy interkaláris szegmens és három állkapocsszegmens. Ezután három mellkasi és tíz hasi szakasz, valamint egy anális lebeny alakul ki.
Sok rovarnál az embrió három szakaszon megy keresztül, amelyeket a végtag primordiumainak eltérő összetétele jellemez: protopod, polypod és oligopod (339. ábra).
A rovarok embrionális fejlődését a blasztokinézis jelensége jellemzi. Ez egy változás az embrió testének helyzetében a tojásban, amelyben a sárgája tartalékok a legteljesebben hasznosulnak.
A rovarok blasztokinézisének két típusát írja le A. G. Sharov. A nem teljes átalakulású rovaroknál az embrió eleinte háttal felfelé és fejjel a pete elülső vége felé helyezkedik el, majd a magzatüreg kialakulásakor az embrió hasi oldalával felfelé fordul, és a feje illetve a tojás hátuljában van.
A blastokinézis a legtöbb teljes átalakulású rovarban és az orthopterában eltérően fordul elő, amikor is az embrió a tojássárgájába süllyed anélkül, hogy a test helyzete megváltozna a tojásban.
A rovarok embrionális fejlődésében megnyilvánulnak a szárazföldi élethez való alkalmazkodás: védőmembránok (chorion, serosa, amnion), tápanyagellátás (sok sárgája), folyadékkal teli magzatvíz.
A kikelés előtt a kialakult rovarlárva folyadékot nyel le a magzatvíz üregéből, aminek következtében a test turgora megnő. A lárva a fejével áttöri a choriont, amelyen gyakran tojásfogak vagy tüske található.
Posztembrionális fejlődés. A rovarok posztembrionális fejlődésének időszakában, a tojásból való kikelés után a fiatal állat növekedése egymást követő vedléseken és minőségileg eltérő fejlődési fázisokon keresztül történik. Az ontogenezis vagy egyedfejlődés során a rovarok 3-4-30-szor vedlenek. A linkek száma átlagosan 5-6. A vedlés közötti intervallumot szakasznak, a fejlődési állapotot pedig életkornak nevezzük. A lárvától a felnőttig terjedő fejlődés során bekövetkező morfológiai változásokat metamorfózisnak nevezzük. Az alsó szárnyatlan formák kivételével minden rovarnál a kifejlett állapot elérése után az imágók növekedése és vedlése megszűnik. Ezért például az azonos fajhoz tartozó bogarak méretének eltérései nem tulajdoníthatók különböző korcsoportoknak, hanem csak az egyéni változatosság megnyilvánulásának tekintendők.
A rovarok posztembrionális fejlődésének három fő típusa van: 1) közvetlen fejlődés metamorfózis nélkül – ametabolizmus vagy protometabolizmus; 2) fejlődés hiányos átalakulással vagy fokozatos metamorfózissal, - hemimetabolizmus; 3) fejlesztés teljes átalakulással, i.e. kifejezett metamorfózissal - holometabolizmussal.
Ametabolizmus, vagy közvetlen fejlődés, csak a sörtefarkú rendbe (Thysanura) tartozó elsődleges szárnyatlan rovaroknál figyelhető meg, amelyek közé tartozik a közönséges ezüsthal (Lepisma). Ugyanilyen típusú fejlődés figyelhető meg Entognathában: rugófarkú (Collembola) és kétfarkú (Diplura).
Az anyagcsere következtében a tojásból egy kifejletthez hasonló lárva jön ki. A különbségek csak a méretre, a test arányaira és az ivarmirigyek fejlettségi fokára vonatkoznak. A szárnyas rovarokkal ellentétben még képzeletbeli állapotban is vedlik.
Hemimetabolizmus- nem teljes átalakulás, vagy fokozatos metamorfózissal járó fejlődés. Sok szárnyas rovarra jellemző, például csótányokra, szöcskékre, sáskákra, poloskákra, kabócákra stb.
A hemimetabolizmussal a tojásból egy lárva jön ki, hasonlóan a kifejlettekhez, de kezdetleges szárnyakkal és fejletlen ivarmirigyekkel. Az ilyen, kifejlett, szárnyakkal rendelkező lárvákat nimfáknak nevezik. Ezt a nevet az ókori görög mitológiából kölcsönözték, és lányok formájú isteni szárnyas lényekre utal. A rovarnimfák többször vedlenek, és minden vedléssel megnő a szárnyuk kezdetlegessége. Egy idősebb nimfa vedlik, és egy szárnyas imágó bújik elő belőle. A 340. ábra a sáskafejlődés fázisait (tojások, 1-5. lárvák nimfái és imágók) mutatja be a hiányos átalakulás példájaként. Ezt a tipikus nem teljes átalakulást hemimetamorfózisnak nevezik.
A hiányos metamorfózisú rovarok között vannak olyan fejlődési esetek, amikor a nimfák észrevehetően különböznek a felnőttektől a speciális lárva-adaptációk - ideiglenes szervek - jelenlétében. Ilyen fejlődés figyelhető meg a szitakötőkben, májusban, kőlegyekben. Ezeknek a rovaroknak a nimfái vízben élnek, ezért hívják őket

Rizs. 340. Fejlődés hiányos metamorfózissal a Locusta migratoria sáskában (Holodkovszkij szerint): 1 - prothorax, 2 - mesothorax szárnyrügyekkel, 3 - metathorax szárnyrügyekkel
naiádok (vízi nimfák). Ideiglenes szerveik, például légcsőkopoltyúik vannak, amelyek szárazföldi felnőtteknél eltűnnek. És a szitakötő lárváinak még mindig van "maszkja" - egy módosított alsó ajak, amely a zsákmány megragadására szolgál.
Holometabolizmus- teljes átalakulás. A fejlődési fázisok a holometabolyban: tojás - lárva - báb - imágó (341. ábra). Ez a fejlődés a bogarakra, a lepkékre, a kétszárnyúakra, a szűzhártyákra, a szúnyogokra és a csipkefélékre jellemző.

Rizs. 341. Fejlődés teljes átalakulással a selyemhernyó Bombyx csúcsában (Leines szerint): A - hím, B - nőstény, C - hernyó, D - gubó, E - gubó báb
A teljes metamorfózisú rovarlárvák nem hasonlítanak a kifejlettekre, és gyakran ökológiailag különböznek egymástól. Így például a májusi bogarak lárvái a talajban élnek, az imágók pedig a fákon. Sok légy lárvája a talajban, a rothadó aljzatban fejlődik, a kifejlett kifejlettek pedig repülnek és meglátogatják a virágokat, nektárral táplálkoznak. Az ilyen rovarok lárvái többször vedlenek, majd bábokká alakulnak. A bábfázisban hisztolízis következik be - a lárvaszervek elpusztulása és a hisztogenezis - egy felnőtt rovar szervezetének kialakulása. A bábból egy szárnyas rovar bújik elő – egy imágó.
Így a rovaroknál a következő típusú posztembrionális fejlődés figyelhető meg: ametabolizmus vagy protomorfózis (tojás - lárva (hasonlóan az imago-hoz) - imago); hemimetabolia - hiányos átalakulás (tojás - nimfa - imago): hemimetamorphosis - tipikus változat, hypomorphosis - csökkent metamorfózis, hipermorfózis - fokozott metamorfózis; holometaboly - teljes átalakulás (tojás - lárva - báb - felnőtt): holometamorfózis - tipikus változat, hipermetamorfózis - többféle lárvával.
A rovarlárvák típusai teljes metamorfózissal. A holometabolikus rovarok lárvái egyszerűbb szerkezetűek, mint a felnőttek. Nincs összetett szemük, szárnyuk kezdete; a szájkészülék rágcsáló típusú, az antennák és a lábak rövidek. A végtagok fejlődése szerint a lárvák négy típusát különböztetjük meg: protopodák, oligopodák, polipodák és apodák(342. ábra). A protopodák lárvái a méhekre, darazsakra jellemzőek. Csak a mellkasi lábak kezdetei vannak. Ezek a lárvák inaktívak, fésűben fejlődnek a dolgozók gondos kezelésével. Az oligopod lárvák gyakoribbak, mint mások, három pár gyaloglábak normális fejlődése jellemzi őket. Az oligopodák közé tartoznak a bogarak lárvái, a fűzők. A többlábú lárvák vagy hernyók három pár mellkasi lábon kívül több pár műlábat is tartalmaznak a hason. A ventrális crura a hasüreg kiemelkedéseit jelenti
testfalak és medvekampók és tüskék a talpon. A hernyók a lepkékre és a fűrészlegyekre jellemzőek, az apodikus, vagy lábatlan lárvák a kétszárnyúak rendjében, valamint egyes bogaraknál (márna lárvái, fúrólyuk), lepkéknél figyelhetők meg.

Rizs. 342. Teljes metamorfózisú rovarlárvák (Barnes-ból): A - protopod, B, C - oligopod, D - polipod, E, F, G - apod
A mozgási módszerek szerint a teljes metamorfózissal rendelkező rovarlárvákat felosztják campodeoid hosszú, rugalmas testtel, futó lábakkal és cerci érzéssel, erucoid húsos, enyhén ívelt testtel, végtagokkal vagy anélkül, huzal- merev testtel, kerek átmérőjű, alátámasztó cercikkel - urogomphs ill féregszerű- lábatlan.
A Campodeoid lárvák számos ragadozó bogara jellemzőek - földi bogarak, sárbogarak. A talajban lévő lyukakon keresztül mozognak. Tipikus erucoid lárva a májusi bogár lárvája, trágyabogár, bronz. Ezek üreges lárvák. A drótférgek a csattanó- és sötétbogarakra jellemzőek, amelyek lárvái aktívan áthaladnak a talajban. Számos féregszerű lárva. A talajban, a növényi szövetekben mozognak. Ide tartoznak nemcsak a Diptera lárvák, hanem egyes bogarak, lepkék, fűrészlegyek is, amelyek például a növényi szövetekben fejlődnek.
bábfajták. A bábok szabadok, fedettek és rejtettek (343. ábra). A szabad báboknál a szárnyak és a végtagok kezdetlegessége jól látható és szabadon elkülönül a testtől, például a bogaraknál. A fedett báboknál minden alapelem szorosan a testhez tapad, például a pillangóknál. A szabad bábok bőre vékony, puha, míg a fedett báboké erősen szklerotizálódott.

Rizs. 343. A bábok fajtái rovarokban (Webertől): A - szabad bogár, B - fedett lepke, C - rejtett légy; 1 - antenna, 2 - szárnyak alapjai, 3 - láb, 4 - spirálok
lárva bőre, amely hamis gubót képez - puparia. A bábházban egy nyitott báb található. Ezért a rejtett chrysalis csak egy változata a szabadnak. A pupariák sok légyre jellemzőek.
Gyakran az utolsó lárva lárvája gubót sző a bábozás előtt. Így például a selyemhernyó-lepkék hernyója selymet választ ki a selyemmirigyekből, amelyből sűrű gubót fon. Egy ilyen gubó belsejében egy fedett krizális található. Egyes hymenoptera - hangyákban, valamint a fűzőben a gubó belsejében nyitott vagy szabad báb található. A fűzős lárvákban, mint például az aranyszemek, a gubófonalakat a malpighi erek állítják elő, és a végbélnyílásból ürülnek ki.
A metamorfózis élettana. A metamorfózis folyamatában két egymással összefüggő folyamat megy végbe: a hisztolízis és a hisztogenezis. A hisztolízis a lárvaszervek szöveteinek lebontása, a hisztogenezis pedig egy felnőtt rovar szerveinek kialakulása. A nem teljes metamorfózisú rovaroknál ezek a folyamatok fokozatosan mennek végbe a nimfa fázisban, a teljes metamorfózisú rovaroknál pedig a bábfázisban.
A hisztolízis a fagociták és enzimek aktivitása miatt következik be. Ebben az esetben mindenekelőtt a zsírtest, a lárvaizmok és néhány más szerv pusztul el, amelyek a fejlődő szövetek által fogyasztott tápanyag-szubsztrátummá alakulnak.
A hisztogenezis vagy egy felnőtt rovar szerveinek kialakulása elsősorban a képzeletbeli korongok - differenciálatlan sejtekből származó kezdetek - fejlődése miatt következik be. A képzeletbeli korongok a lárvafázisban, sőt az embriogenezis során is lerakódnak, és belső kezdetlegességet képviselnek. A képzeletbeli lemezek fejlesztik a szemet, a szárnyakat, a szájkészüléket, a lábakat, valamint a belső szerveket: izmokat, ivarmirigyeket. Az emésztőrendszer, a malpighi erek és a légcső nem pusztul el, de erősen differenciálódnak a metamorfózis folyamatában. A legkevésbé metamorfizált szív és idegrendszer. Az idegrendszer metamorfózisa során azonban gyakran megfigyelhető a ganglionok oligomerizációja (fúziója).
A metamorfózis folyamatát a belső elválasztású mirigyek irányítják (329. ábra). neuroszekréciós sejtek az agy hormonokat választ ki, amelyek serkentik az aktivitást szívtestek, melynek hormonjai a hemolimfán keresztül serkentik prothoracalis vedlési hormont termelő (prothoracalis) mirigyek - ekdizon. Az ekdizon elősegíti a vedlési folyamatot: a régi kutikula részleges feloldódását és hámlását, valamint egy új képződését.
A metamorfózis folyamatában a tevékenység szomszédos testek termelő fiatalkori hormon. Nagy koncentrációban a lárva vedlése lárva képződéséhez vezet
következő kor. A lárvák növekedésével a szomszédos testek aktivitása gyengül, a juvenilis hormon koncentrációja csökken, a prothoracalis mirigyek fokozatosan degenerálódnak. Ez oda vezet, hogy a lárvák a bábfázisba, majd a kifejlettbe olvadnak.
A szomszédos testek mesterséges átültetése például egy utolsó korú sáska nimfába hozzájárul ahhoz, hogy az nem a kifejlett fázisba olvad, hanem egy nagyobb, további korú lárvává. Felnőtt fázisban a juvenilis hormon szabályozza az ivarmirigyek fejlődését, az ekdizon hormon a prothoracalis mirigyek csökkenése miatt már nem termelődik.
A metamorfózis eredete. Számos hipotézis létezik a rovarok metamorfózisának eredetéről. Hosszú ideig viták voltak arról, hogy mely rovarok fejlettebbek evolúciósan - teljes vagy hiányos átalakulással. Egyrészt a hiányos metamorfózisú rovarnimfák progresszívebben fejlődnek, mint a teljes metamorfózisú rovarlárvák, másrészt az utóbbiak előrehaladott bábfázisúak.
Jelenleg ezt az ellentmondást G. S. Gilyarov, A. A. Zakhvatkin és A. G. Sharov metamorfózis eredetének hipotézise hárította el. E hipotézis szerint a rovarokban a metamorfózis mindkét formája egy egyszerűbb fejlődési típustól függetlenül fejlődött ki - a protomorfózistól, amelyet az elsődleges szárnyatlan rovaroknál figyeltek meg, például a sörtefarkú rovaroknál (Thysanura).
Protomorfózis esetén a fejlődés közvetlen, sok vedlés figyelhető meg a lárvafázisban, majd a képzeletbeli állapotban. E rovarok fejlődésének minden fázisa ugyanabban a környezetben történik.
Feltételezik, hogy az evolúció során a rovarok a felső talajrétegben lévő félig rejtett létből a felszínen és a növényeken éltek át. Ezt az új élőhelyre való átmenetet egy jelentős aromorfózis koronázta meg - a szárnyak fejlődése és a repülés.
A nyílt élőhelyek fejlődése a rovarok egyedfejlődésének alakulásában is megmutatkozott. A rovarok ontogenezisének evolúciója láthatóan két fő irányt követett.
Egy esetben a fejlődés embrionizálódási folyamata zajlott, ami a fejlődés későbbi fázisaiban a sárgájában gazdag tojásokból a rovarok kikeléséhez vezetett. Ez oda vezetett, hogy a lárvákat nimfák képződésével képzelték el. Így fejlődtek ki a nem teljes metamorfózisú rovarok. Ez az evolúciós út vezetett a lárvák fokozatos fejlődéséhez, amelyek hasonló életmódot folytatnak a felnőttekkel.
A másik esetben éppen ellenkezőleg, a fejlődés deembryonizációs folyamata ment végbe, vagyis a sárgájában szegény tojások a fejlődés korábbi szakaszaiban keltek ki. Ez a rovarlárvák és az imágók morfoökológiai eltéréséhez vezetett. A lárvák leegyszerűsödtek és alkalmazkodtak
védettebb környezetben élni, a táplálkozási funkciót ellátva, a felnőttek pedig elsősorban a szaporodási és letelepedési funkciót kezdték el ellátni. A teljes átalakulással járó rovarlárvák fejlődésének deembrionizálása mellett számos ideiglenes alkalmazkodást fejlesztettek ki a különféle létfeltételekhez. Így amfigenezis (divergencia) következett be a lárvák és az imágók evolúciójában a teljes metamorfózissal járó rovaroknál. A lárvák és imágók amfigenezise a morfológiai adaptációk szempontjából igen mélynek bizonyult, ami komoly ellentmondásokat szült az ontogenezisben. Sikeresen megoldották őket a bábfázis megjelenése, amely során a lárvaszervezet radikális átstrukturálása az imaginálisra. Ez lehetővé tette a teljesen átalakult rovarok számára, hogy az ökológiai rések szélesebb körét elsajátítsák, és példátlan virágzást érjenek el a Földön élő állatok között.
Rovartenyésztés. A legtöbb rovarra a kétivarú ivaros szaporodás a jellemző. Sok faj nemi dimorfizmust mutat. Például a hím szarvasbogarak mandibulája szarvra módosult, a hím orrszarvúbogarak fején szarv, a pronotumán pedig púp található. Ez e fajok párzási viselkedésének köszönhető, amelyet a hímek nőstényért való küzdelme kísér. A nemek közötti kapcsolat a különböző fajoknál rendkívül változatos. A Dolichopodidae családba tartozó hím kétszárnyúak "ajándékot" hoznak a nősténynek - egy elkapott legyet, és lábtükrökkel táncolnak. A nőstény imádkozó sáskák ragadozók, és párzás közben megeszik a hímet.
A legtöbb rovar tojást rak, de gyakran megfigyelhető az élve születés is. Ebben az esetben a peték a női nemi szervekben fejlődnek ki, és lárvákat szül. Például a blowflies-szarkofágok (Sarcophagidae) élő lárvákat raknak a húsra, amelyek fejlődése nagyon gyorsan megy végbe. Nem csoda, hogy az ókorban azt hitték, hogy a férgek a húsban spontán módon keletkeznek. Lehetséges, hogy nem veszi észre, ahogy egy légy meglátogatta a nyíltan fekvő húst, és hirtelen felfedezi a hirtelen megjelent fehér lárvákat.
Az életképes fajok közé tartozik a birkavérszívó légy is, néhány barlangban élő bogarak.
A kétivarú ivaros szaporodás mellett számos rovar partenogenezist mutat – megtermékenyítés nélkül fejlődik. A rovarok különböző rendjéből számos faj létezik, amelyekre a partenogenezis jellemző. A partenogenezis lehet kötelező – kötelező, akkor a faj összes egyede csak nőstény. Tehát alpesi viszonyok között, északon és más kedvezőtlen körülmények között partenogenetikus bogarak, orthoptera, füles és fűzős bogarak találhatók. A partenogenezis a kétivarú fajoknál is előfordul, amikor a peték egy részét megtermékenyítve, egy részét megtermékenyítés nélkül rakják le. Például a méhlepények megtermékenyítetlen tojásokból fejlődnek ki.
Hasonló partenogenezis megtalálható más Hymenoptera (hangyák, fűrészlegyek), termeszek, egyes poloskák és bogarak esetében is. És például a levéltetvek életciklusában generációváltás történik: biszexuális és partenogenetikus. Egyes esetekben a partenogenezis opcionális (átmeneti) lehet, amely csak kedvezőtlen körülmények között nyilvánul meg. A rovarok partenogenezise hozzájárul a magas populációk fenntartásához.
A partenogenezis egyik változata a pedogenezis - szaporodás megtermékenyítés nélkül a lárvafejlődési fázisban. Ez a rovarok speciális fejlődési módja, amikor az ivarmirigyek érése megelőzi a többi szervet. Például egyes epeszúnyogfajok lárvafázisban szaporodnak. Az idősebb lárvák fiatalabb lárvákat szülnek. Pedogenezist figyeltek meg az egyik bogárfajnál, amelynek lárvái részben tojásokat raknak, részben pedig lárvákat szülnek. A pedogenezis a kétivarú szaporodás mellett növeli a faj egyedszámát.
A rovarok életciklusai. Ellentétben az ontogenezissel vagy a rovarok egyedfejlődésével, az életciklus egy faj fejlődése, amely rendszerint többféle ontogenezist is magában foglal. Az ontogenetika egy egyed életére korlátozódik a tojástól a pubertás kezdetéig, majd a természetes halálig. Az életciklus egy faj folyamatos fejlődésének visszatérő része. Így a rovaroknál a legjellemzőbb esetben az életciklus két konjugált és morfofiziológiailag eltérő ontogenetikus hím és nőstény egyedből áll, amelyek ivarosan szaporodnak és saját fajtájukat szaporítják. A partenogenetikus fajoknál pedig az életciklust csak egyfajta női ontogenetika jellemzi.
A rovarok életciklusai sokfélék a szaporodási típusok, a generációk összetétele és váltakozása tekintetében. A rovarok életciklusának következő típusai különböztethetők meg.
1. Életciklusok nemzedékek váltakozása nélkül biivarú ivaros szaporodással. Ez az életciklus leggyakoribb típusa, amely a csak hímekből és ivarosan szaporodó nőstényekből álló dimorf fajokra jellemző. A legtöbb bogarak, lepkék, poloskák ciklusai ilyenek.
2. Életciklusok generációk váltakozása nélkül, partenogenetikus szaporodással. Az ilyen fajok monomorfok, csak partenogenetikus nőstényekből állnak, amelyek megtermékenyítés nélkül rakják le a tojásokat.
A partenogenetikus fajok különösen gyakoriak a levéltetvek, psyllidák és más homopterák körében. A partenogenetikus bogarak, poloskák, szöcskék és kokcidák gyakoriak a magashegyi körülmények között.
3. A rovarok életciklusának legritkább típusa a generációk váltakozása nélküli ciklus a hermafrodita fajok ivaros szaporodásával.
Ismert egy amerikai légyfaj, amely csak hermafrodita egyedekből áll. A fejlődés korai szakaszában az imágók hímként, a későbbi szakaszokban nőstényként működnek. Ezért minden egyed tojik, ami növeli a faj egyedszámát.
4. Életciklusok generációváltás nélkül az ivaros szaporodással és fakultatív partenogenezissel polimorf fajokban, például társas rovarokban. A faj ivaros egyedekből - hímek és nőstények, valamint termékeny - dolgozó egyedekből áll, amelyek nem vesznek részt a szaporodásban. E fajok közé tartoznak a méhek, hangyák és termeszek. Az ilyen életciklusokat bonyolítja, hogy a nőstények a megtermékenyített peték mellett partenogenetikusakat is raknak, amelyekből például a méhek haploid hímeket - drónokat, a megtermékenyítettekből pedig nőstényeket és nőstényeket - fejlesztenek. Egyes lovasok, tripszek, kokcidák hasonlóan fejlődnek.
Más fajoknál a fakultatív partenogenezis másképpen nyilvánul meg: nem a hímek, hanem a nőstények fejlődnek ki megtermékenyítetlen petékből. De ebben az esetben a diploid kromoszómák a nőkben a haploid magok fúziójával helyreállnak. Egyes botrovaroknál, sáskáknál, fűrészlegyeknél, kokcidáknál ismert ilyen fejlődés.
5. Életciklusok a szexuális generáció és a partenogenetikus (heterogónia) váltakozásával. Sok levéltetűben és filoxérában a szárnyas hímek és nőstények ivaros nemzedékén kívül a partenogenetikus nőstények több generációja is létezik, amelyek szárnyasak vagy szárnyatlanok.
6. Életciklusok ivaros generáció váltakozásával és több generáció pedogenezisével. Például egyes epeszúnyogoknál az ivaros szaporodás után, amelyben hímek és nőstények vesznek részt, a lárvák partenogenetikus szaporodása (pedogenezis) következik be. A tenyészlárvák több generációja után, amelyek saját fajtájuk élve születése után elpusztulnak, a lárvák utolsó generációja bebábozódik, és szárnyas nőstényeket és hímeket hoz létre.
7. Életciklusok a szexuális generációk (férfiak és nőstények) és aszexuálisok váltakozásával. Az ivaros szaporodás után a nőstények megtermékenyített petéket raknak, amelyek poliembriónián mennek keresztül. Ez ivartalan szaporodás az embrionális fázisban. A tojás összezúzódik, és a morula fázisban lévő embrió bimbózás útján szaporodni kezd. Egy tojásból több tucat embrió képződhet. Ilyen
Így az életciklusok osztályozása a következőképpen ábrázolható.
ÉN. Nincs generációváltás:
- 1) kétivarú ivaros szaporodás esetén (május bogár);
- 2) partenogenetikus szaporodással (alpesi bogarak, szöcskék);
- 3) hermafrodita egyedek ivaros szaporodásával (amerikai légy);
- 4) polimorf fajok (méhek) ivaros szaporodásával és részleges partenogenezisével.
II. Nemzedékek váltakozásával:
- 1) heterogónia: az ivaros nemzedék és számos partenogenetikus (levéltetvek, filoxéra) váltakozása;
- 2) heterogónia: az ivaros nemzedék és több pedogenetikai nemzedék váltakozása (néhány epehólyag);
- 3) metagenezis: a szexuális generáció váltakozása a poliembrióniával (lovasok).
A rovarok szezonális ciklusai. Ha az életciklus alatt egy faj morfogenezisének ciklikusan ismétlődő részét a fejlődés egyik fázisától ugyanabba a fázisig értjük, akkor az évszakos fejlődési cikluson egy faj egy éves évszakok alatti fejlődésének jellemzőjét értjük. téltől télig).
Így például a májusi bogár életciklusa 4-5 évig tart (a tojástól a kifejlett egyedekig), és e faj szezonális ciklusát az a tény jellemzi, hogy az áttelelt lárvák tavasszal bebábozódnak, és a fiatal bogarak szaporodnak. Nyáron, ősszel és télen különböző korú lárváik találhatók. Az év során kifejlődő generációk számát voltságnak nevezzük.
Vannak olyan fajok, amelyek évente több generációt adnak. Ezek többvoltaikus fajok. Például egy házilégy szezononként 2-3 generációt tud adni, és a felnőtt fázisban hibernálódik. A legtöbb rovar egyvoltos, évente egy generációt hoz létre.
A természetben a rovarok szezonális ciklusait a különböző fejlődési fázisok naptári időszakai jellemzik. A fajok szezonális ciklusainak fontos jellemzői az aktív életük időzítése és a diapauza (átmeneti késleltetés a fejlődésben) télen vagy nyáron. A fajok életciklusának a helyi szezonális jelenségeknek megfelelő szabályozását a környezeti tényezők és a szervezet neurohumorális rendszere biztosítják.
A munkásméh, méh, drón embrionális, vagy embrionális fejlődése magában foglalja mindazokat a változásokat, amelyek a tojáshéj alatt végbemennek, aminek következtében egy egysejtű tojásból többsejtű lény, a lárva jön létre (32. kép). ). A méh által éppen lerakott tojást általában egyik végével a sejt aljára ragasztják, és függőlegesen áll rajta. A tojás hossza 1,6-1,8 mm, szélessége 0,31-0,33 mm. Enyhén ívelt, hosszúkás henger alakú, a rögzítési helytől ellentétes vége kissé kitágult. Tovább
- A 3. napon a tojás ferde helyzetbe kerül, a 3. napon pedig a sejt alján fekszik. A petesejt szabadon kitágult végén van egy apró nyílás, amelyen keresztül a spermiumok belépnek a méhedényből, miközben a petesejt áthalad a petevezetéken. Ezt a lyukat mikropilának nevezik. Más esetekben a mikropila mind a baktériumok, mind a vírusok számára átjárhatatlan. A tojásnak elülső és hátsó vége van. A lárva feje a tojás szabad elülső végén fejlődik ki.
Az embrionális fejlődés a mag feldarabolásával kezdődik. Az osztódás eredményeként kialakuló leánymagok először véletlenszerűen helyezkednek el a tojássárgájában, majd a citoplazma kis területeivel a tojás perifériájára vándorolnak, és egy folytonos sejtréteget alkotnak - a blastodermát, amely a teljes tojáshéjat kibéleli. a belseje. A tojás domború oldalán a blastoderma sejtek gyorsabban kezdenek növekedni és szaporodni, mint a homorú oldalon lévő sejtek. Ennek eredményeként egy embrionális csík képződik széles szál formájában. Ez az embrió kialakulásának kezdete. A csíracsík a sejtosztódás és differenciálódás, a szövetképződés miatt nő. Ezen folyamatok eredményeként megtörténik a lárva fejlődése. A 2. nap második felében megkezdődik az embrió szegmentálódási folyamata. A csíracsík elülső végén preorális lebeny (akron) képződik, amelyen gumó képződik - a felső ajak rudimentuma. E gumó alatt egy mélyedés jelenik meg - a jövő szája. Ezután - az antennák alapjai. A mellkas és a has szegmentumai hátrafelé válnak le a fejrészről. Ezeknek a szegmenseknek a ventrális (ventrális) oldaláról kinövések jelennek meg - a páros végtagok alapjai. Először a mellkason, később a hason jelentkezik. Ugyanakkor a fejrészben egy határozott tagolódás található: a fej öt szegmensének kezdetlegessége alakul ki. Ezenkívül légzési lyukak jelennek meg - stigmák, valamint a forgó mirigyek és a malpighi erek kezdetei. Ahogy a test és a végtagok szegmensei kialakulnak, az egyes részek szétválnak.
Ezt követően a csíracsík mérete megnő. A blastoderma sejtek szaporodása a csíracsík középső részének invaginációjához vezet (a gastruláció folyamata), amely mélyebbre ereszkedik a tojásba, és elválik a blastodermától. A gasztruláció során először egy hosszanti barázda képződik a csíracsík felületén, majd a barázda külső szélei egymás felé kezdenek növekedni. Ennek eredményeként a későbbi második csíraréteg behatol a csíracsík belsejébe. Amikor a horony szélei összezáródnak, kialakul a külső réteg (ektoderma). Alatta fekszik a belső réteg (mezoderma). A mezodermális réteg végein invaginációk jelennek meg, amelyek sejtcsoportokból állnak, amelyek a harmadik csíraréteget (endodermát) alkotják. Az egyes szervek kezdetlegességei kezdenek megjelenni. Fejlődésük és differenciálódásuk addig tart, amíg a lárva ki nem kel a sejtből.
Az ektodermából a testfal, az elülső és hátsó belek, a légcső és az idegrendszer, valamint számos mirigy képződik; az endodermából - a csípés kezdetei, a szárnyak és a végtagok, a genitális traktus végszakaszok, az idegrendszer és az érzékszervek; a mezodermából - izmok, zsírtest, nemi mirigyek.
Egy teljesen kialakult embrió, amely lényegében lárvává alakult, kitölti az egész tojást. Elkezd intenzíven mozogni, levegőt szív a légcsőbe, lenyeli a magzatvizet, ezáltal növeli testének térfogatát. A lárva kiszabadul az embrionális membránokból, áttöri a choriont (a tojás másodlagos héját, amely a tüszőhámsejtek átalakulásával alakul ki) és kikel, vagyis kijön. A mézelő méhek embrionális fejlődése általában körülbelül 3 napig tart. A hőmérséklet változása növelheti vagy lerövidítheti az embrionális fejlődési időszakot.
A lárva tojásból való kiszabadulása után megindul a posztembrionális fejlődés, amelyet metamorfózis (transzformáció) kísér.
A metamorfózis a szervezet szerkezetének mélyreható átalakulása, melynek eredményeként a lárva kifejlett rovarrá alakul. A rovarok posztembrionális fejlődésének természetétől függően a metamorfózis két típusát különböztetjük meg:
hiányos (hemimetabolizmus), amikor a rovar fejlődését csak három szakasz - tojások, lárvák és a felnőtt fázis (felnőtt) áthaladása jellemzi;
teljes (holometabolizmus), amikor a lárva átmenete a felnőtt formába egy köztes szakaszban - a bábállapotban - történik.
Hiányos átalakulással a korai szakaszban élők és a felnőttek életmódja hasonló. Az ilyen kifejlődésű lárva egy kifejlett rovarhoz hasonlít, és hozzá hasonlóan összetett szemei, szájszervei, majd kívülről jól látható szárnyaik is vannak. A szárnyak rudimentumai minden egyes következő vedléskor megnövekednek, és az utolsó preimaginális korban már a has több szegmensét is befedhetik. Az utolsó preimaginális korú rovarban az imaginális struktúrák teljesen kifejlődnek, és az utolsó teljesen kialakult vedlés eredményeként egy imágó jelenik meg. A hiányos átalakulás jellemző az orthoptera, poloska, homoptera stb.
Teljes metamorfózis esetén a teljes fejlődési ciklust a tojás, a lárva, a prepupa, a báb és a kifejlett stádiumok áthaladása jellemzi (3. táblázat).
- A méhcsalád egyedeinek fejlődési szakaszainak időtartama, napok
A lárvából a kifejlettbe való átmenet a bábállapotban történik. Ez egy nem tápláló szakasz, amelynek során a hozzá tartozó szövetek
A korábbi szakaszokban képzeletbeli struktúrák alakulnak ki. A legtöbb rovarnál ez a szakasz mozdulatlan. A báb nem reagál a külső ingerekre, vastag takaróba van öltözve. Ennek a szakasznak a végén a héj szétreped, és megjelenik egy felnőtt rovar.
A rovaroknál a metamorfózis szabályozását az endokrin mirigyek (endokrin mirigyek) által termelt hormonok végzik. Jelenleg három metamorfózishormon ismeretes, amelyek szabályozzák a posztembrionális fejlődést: a juvenilis hormon, a vedlési hormon (lárva) és az aktivációs hormon. A juvenilis hormont a szomszédos testekben termelik és tárolják, amelyek két kis sejtszerkezet, amelyek az agy mögött, a nyelőcső oldalain helyezkednek el.
Ha egy fiatal lárváról eltávolítjuk a szomszédos testeket, akkor a következő vedlést követi a bábozás, annak ellenére, hogy normál állapotában még több vedlést kellene végeznie. Ez a hormon határozza meg a lárvák növekedését, szabályozza számos szerv és folyamat fejlődését. Például a zsírtest nem növekszik és nem működik e hormon hiányában.
A vedlési hormon, vagyis az ekdizon a prothoracalis mirigyekben termelődik, amelyek a lárva prothoraxában helyezkednek el az első ideg ganglion felett, az első spiracle közelében. A rovar testében szintetizálódik belőle. koleszterin. A hormon beindítja a vedlési folyamatot, és így közvetve szabályozza a növekedést és a szerkezet kialakulását. Ennek a hormonnak a rovarokba való bejutása duzzanatokat okoz az óriási kromoszómákon. Ezeknek a duzzanatoknak az elemzése kimutatta, hogy intenzív RNS képződési folyamat megy végbe bennük. Úgy gondolják, hogy az ekdizon hatásának első eredménye a gének aktiválása, majd az RNS bioszintézis stimulálása és a megfelelő enzimek kialakulása, amelyek biztosítják a metamorfózis folyamatát.
Az aktiváló hormon speciális neuroszekréciós sejtekben termelődik, amelyek a supraesophagealis ganglion elülső dorzális részében helyezkednek el. Ezek a mirigyek jobban fejlettek a munkásméheknél, kevésbé a királynőknél, és még kevésbé a drónoknál. Az aktiváló hormon befolyásolja a rovarok tevékenységének újraindulását minden vedlés után, és serkenti két másik hormon – az ekdizon és a fiatalkori – képződését.
A mézelő méh teljes metamorfózisú rovar (33. ábra).
Lárva állapot. Intenzív táplálkozás és növekedés jellemzi. A munkásméh-lárva stádiuma 6 napig tart, a királyné - 5, a dróné - 7. Az első napokban a mézelő méhek lárvái annyi táplálékot kapnak, hogy a felszínén úsznak. A takarmány alapja a hipofarin titka.
| 1 g | 3 | 4 5 | 6 7 8 amper; | 10 | 11 12 | 13 14 15 16 17 18 19 20 | 21 |
| w |
Nap
Rizs. 33. A méhfejlődés szakaszai
dajkaméhek geal mirigyei - áttetsző fehér folyadék. A dajkaméhek mirigyei által kiválasztott tej rendkívül tápláló. Szénhidrátokat, fehérjéket, zsírokat, ásványi anyagokat, B csoportba tartozó vitaminokat tartalmaz. Mielőtt táplálékot adna a lárvának, a munkásméh, fejét a sejtbe hajtva, többször kinyitja és bezárja a felső állkapcsokat. Ezután pofájával megérinti a sejt alját, és folyékony táplálékot bocsát ki. Más méhek ápolók ugyanígy adják hozzá a következő adag tejet a lárvát tartalmazó sejthez. A teljes lárvaállapotban körülbelül 10 ezer látogatás történik egy lárvával rendelkező sejtnél.
A fiatal lárvák és az idősebb lárvák tápláléka nem ugyanaz. A második életszakaszban a munkásméh lárvája zabkását kezd kapni - egy keveréket, amelyet a dajkaméhek mézből és virágporból állítanak elő.
A lárvaállapot során a munkásméh-lárvák lineáris méretei több mint 10-szeresére nőnek. Így az újonnan kikelt lárva hossza 1,6 mm, egy 1 naposé 2,6 mm, egy 2 naposé 6 mm, a lárvaállapot végére pedig 17 mm (majdnem teljesen elfoglalja a cella alja). A méhlárva hossza a szakasz végére eléri a 26,5 mm-t.
Az intenzív táplálkozás jelentős testtömeg-növekedést biztosít a lárvák számára. A kikelt lárva súlya körülbelül 0,1 mg. Elsőre
- nap alatt a munkásméh lárvájának tömege 45-ször, a drón lárváéé pedig 85-szörösére nő. A stádium végére a dolgozó egyedek lárváinak súlya 1565-szörösére, az anyáké 2926-szorosára nő.
A lárva megjelenésében élesen eltér a kifejlett méhtől (34. ábra). Féregszerű formájú, fehéres színű puha borítású. Teste egy fejből, egy csuklós törzsből és egy anális lebenyből áll.
A lárva feje kicsi, tompakúp alakú, a kúp tövét a fejkapszula alkotja, amelyet hosszanti varrat oszt ketté.
Rizs. 34. A méhlárva külső (G) és belső (1G) szerkezete "\u003e
Rizs. 34. A méhlárva külső (D) és belső (1D) szerkezete:
A - általános nézet; B - kilátás a fej oldaláról; C - a kifejlett lárva helyzete a sejt alján; G - oldalnézet; y - az antennák alapjai; vg - felső ajak; vh - felső állkapocs; opzh - a forgó mirigy megnyitása; ng - alsó ajak; p - száj; ndg - supraesophagealis ganglion; PC - előbél; a - aorta; sk - középső bél; zht - kövér test; zpzh - a nemi mirigy csírája; s - szív; zk - hátsó bél; ms - malpighi edény; bn - hasi ideglánc; pzh - fonóvas; pdg - subesophagealis ganglion
két egyenlő domború részre, az úgynevezett arcokra. A kúp csúcsát a fej elülső részén a felső ajak és más szájrészek alkotják. A fej azon részét, beleértve az alsó ajkát, a felső állkapcsot és a fejkapszula azon részét, amelyhez rögzítik, egy mély horony választja el a fejkapszulától, amely mindkét oldalon a felső állkapocs töve között fut. és a maxilla és a fej hátsó határáig terjed.
A mandibulák a fejkapszulából kúp alakú szervek formájában nyúlnak ki. Az antennális rudimentumok a fejkapszulán helyezkednek el a mandibula alapja felett, kerek gumók formájában.
A mellkasi szakaszokon lévő lárváknak nincs külső eleme sem a szárnyaknak, sem a lábaknak. Nincsenek összetett szemek. A lárva kutikula alatt azonban feltűnnek a kezdetlegességeik, amelyeket képzeletbeli rügyeknek neveznek. A lábak imaginális bimbói orsó alakúak, keresztirányú barázdák boncolják. A szárnyak rudimentumai a hypodermis két lapos, befelé hajlított nyúlványából állnak.A második és harmadik mellkasi szegmens oldalain helyezkednek el.A régióban a hasoldali nyolcadik és kilencedik hasi szakaszból a nemi szervek rudimentumai jelennek meg.
Megjegyzendő, hogy a szárnyak, antennák, szemek és lábak fejlődése nem a redukált lárvastruktúrák átstrukturálásával történik, hanem az embrionális szövetek inaktív állapotban lévő elszigetelt területeiről, úgynevezett rudimentumokról fejlődnek ki.
A lárvák a belső szervek felépítésében különböznek a kifejlett méhektől (lásd 34. ábra). A lárva testében a központi helyet a bélcsatorna foglalja el, amely három részből áll - az elülső, a középső és a hátsó bélből. Az előbél egy rövid vékony csőnek tűnik, a hátsó végén kissé kitágult. Az elülső bélben van egy kis szájüreg, egy rövid garat és a nyelőcső. A kifejlett méhekre jellemző mézes golyva hiányzik. Az előbél falában olyan izmok találhatók, amelyek biztosítják a folyékony takarmány felvételét. A nyelőcső és a középbél találkozásánál egy kis gyűrű alakú redő található, amely szelep szerepét tölti be. Lezárja a nyelőcső lumenét, és megakadályozza, hogy a középbél tartalma visszafolyjon a nyelőcsőbe. A lárvaállapot végére a szelep helyén vastag szeptum képződik, amely befedi a középbelet. 1
A középbél a bél legnagyobb része, és a lárva testének legnagyobb részét foglalja el. Ennek a szakasznak a megkülönböztető jellemzője, hogy nincs kapcsolata a hátsó béllel, aminek következtében a táplálék emésztetlen részei benne maradnak a teljes lárvaállapotban. E két szakasz összekapcsolása csak a gubó fonása előtt történik, amikor a lárva már lezárt. Ebben az időszakban a középső beleket a hátsó béltől elválasztó membrán áttörik, és a széklet a hátsó bélbe kerül, majd a testből a sejt aljába kerül.
A középső bélben a lárvák malpighi erekkel rendelkeznek. A mézelő méhnek négy van belőlük. Ezek hosszú, enyhén csavarodott csövek, amelyek a középső bélben futnak. A lárva élete során a malpighi erek zárva vannak. Csak a lárvaállapot 6. napjára, amikor az ott felhalmozódott bomlástermékektől erősen megduzzadnak, a hátsó végek és a tartalom áttörik, átjutnak a hátsó bélbe, majd eltávolítják. Így a lárva növekedése és táplálkozása során nem ürít.
A hátsó bél kicsi, keskeny cső, hegyesszögben ívelt és a végbélnyílásban végződik.
A lárva szíve 12 azonos szerkezetű kamrából áll (egy felnőtt méhben csak öt van), és egy vékony falú cső, amely közvetlenül a has és a mellkas kutikulája alatt helyezkedik el. A mellkasi régió második szegmensében halad át az aorta. Úgy néz ki, mint egy lefelé hajlított cső, amely azután a fejbe jut, és az agy elülső felületén végződik, ahol a hemolimfa a testüregbe ömlik. Az aorta felszínén a légcsők sűrű hálózata található.
A szív minden kamrája között nyílások vannak, amelyeken keresztül a hemolimfa belép a szívbe. Tekintettel arra, hogy az egyes kamrák vékony elülső végei benyúlnak a következő kamrába, a folyadék fordított áramlása nem megengedett.
A lárva légzőrendszere meglehetősen egyszerű, és két nagy hosszanti légcsőtörzs képviseli a test oldalain, amelyekből kis ágak terjednek az egész testben. A szelvények oldalán 10 pár spirál található, amelyek keskeny kitingyűrűkkel körülvett mélyedések.
Az idegrendszer két egyszerűen elrendezett fej ganglionból (egy nagy supraoesophagealis és egy kis subglottalis) és egy ventrális idegzsinórból áll, amely a mellkas és a has alsó részén húzódik. A hasi lánc 11 ganglionból áll - 3 mellkasi és 8 hasi. Az utolsó hasi ganglion a tizenegyedik szegmens közepén van; az utolsó három szegmens ganglionjainak összeolvadásával jött létre. A ganglionok páros összeköttetésekkel kapcsolódnak egymáshoz. A nyelőcső alatti ganglion két összekötőn keresztül is kapcsolódik a ventrális idegzsinórhoz.
A nemi szervek a munkásméh lárvájában már csecsemőkorban jelen vannak, és a nyolcadik szegmensben elhelyezkedő két gerinc képviseli őket. Hosszúságuk átlagosan 0,27 mm, szélességük 0,14 mm. A teljes lárvaállapot alatt a petefészek-primordiák kialakulása következik be. A 2 napos lárvában jelentéktelen méretűek, a 6 napos lárvában már megnyúltak, és a kilencedik szegmensen keresztül a farokvég felé helyezkednek el. A lárvaállapot végére egy dolgozó méhben a tojáscsövek száma eléri a 130-150-et. A következő szakaszokban, amikor a lárva bábbá változik, a lárvaszövetek szétesnek, és a munkásméhben a petékcsövek száma 20-ról 3-ra csökken, míg a méhben a petefészkek bábállapotban fejlődnek tovább.
A zsírtest jól fejlett a lárvában, ahol a tápanyagok felhalmozódása történik. Idősebb lárváknál a zsírtest eléri a testtömeg 60%-át, és szinte a teljes üreget kitölti a lárva középbéle és kutikulája között. A zsírtest a zsírok, fehérjék és szénhidrátok koncentrációja, amelynek köszönhetően a báb a jövőben fejlődik. A zsírtest a tényleges zsírsejteken kívül kiválasztó sejtekből és enocitákból is áll, amelyek megfogják a húgysavsókat.
A forgó mirigy csak a lárvákban van jelen. Ennek a mirigynek a váladéka anyagként szolgál a gubó fonásához, mielőtt a lárva a bábállapotba kerülne. A forgó mirigy úgy néz ki, mint két hosszú cső, amelyeket a hasi oldalról a középső bél alá helyeznek. Az elülső hasüregben mindkét cső egy páratlan kiválasztó csatornába csatlakozik, amely az alsó ajak hegyén lévő szájnyílás alatt nyílik.
A teljes stádiumban a lárva egy szúrócska kezdetlegességeit fejleszti, amelyek közül egy pár a tizenegyedik, két pár a tizenkettedik szegmensben található. Szegmensenként van köztük a nemi szervnyílás kezdete. A nagy mérgező mirigy rudimentuma csak a lárvaállapot vége felé jelenik meg, a bábállapot elején pedig a kilencedik szegmensen a borítás mélyüléséből kis mérgező mirigy alakul ki.
A lárvák tojásból való kiszabadulása utáni 5-6. napon a méhek viaszkupakkal kezdik lezárni a sejtet. A kupakot alkotó anyag összetétele a viasz mellett pollent, vizet és papírmasszát is tartalmaz. A fedél porózusnak bizonyul, ami miatt a lárva, majd a báb légzéséhez szükséges levegő behatol a sejtbe. Ettől a pillanattól kezdve a lárva befejezi növekedését és abbahagyja a táplálkozást.
Amint a sejt lezáródik, a lárva felegyenesedik benne, a bélfalak összehúzódnak, és az emésztetlen táplálékmaradványok, miután először áttörték a középső bél vékony falát, átjutnak a vastagba, majd onnan kifelé. A lárva ürüléket rak le a sejt egyik sarkában. Ezt követően a lárva gubót pörget, azaz befonja a sejt belső felületét. A gubó fonása során a lárva a forgó mirigy váladékát, a malpighi ereket és a teste falaiból származó ragacsos váladékot használja fel. A fonási folyamatot Velich (1930) tanulmányozta és részletesen leírta.
A gubó pörgetése közben a lárva gyors, ritmikus remegő mozdulatokat végez; a fejtől kiindulva a hullám áthalad a szegmenseken. 1 perc alatt a lárva akár 280 mozdulatot is végrehajt. A remegéssel egyidejűleg a lárva a fej ringatását a test elülső végével hajtja végre. Szaggatott, ellipszis alakú, szabálytalan görbületek fentről lefelé vagy jobbról balra. Ugyanakkor egy átlátszó anyag áramlik ki a nyílásból az alsó ajak szelepével, amelyet a lárva egy vastag vagy több vékony szál formájában választ ki. 2 napon belül a lárva megfordulva megerősíti a gubó falát. A gubó fonása során a dolgozó méhek, drónok és királynők lárváinak viselkedése hasonló volt, a különbség csak a gubó alakjában volt. Ez attól függ, hogy mekkora a sejt, amelyben a lárva található. Például egy munkásméhnél a gubó egy hatszögletű prizma formájának felel meg, melynek alja háromoldalú gúla, a fedél pedig egy lezárt méhsejt pontos lenyomata. A gubót a fiatal méh kilépése után nem távolítják el. Tekintettel arra, hogy egy cellában több tucat generáció jelenik meg, a cella átmérője idővel szűkül. Tehát, ha egy frissen épített fésű normál sejtjének térfogata 0,282 cm3, fenékvastagsága 0,22 mm, akkor a méhek 20 generációja után a sejt kubatúrája 0,248 cm3-re csökken, az alja pedig 1,44 mm-re vastagodik. .
A munkásméh és a királynő lárvája 2 napig, a drón lárvája 3 napig pörget egy gubót. Ezután ötödször vedlik. Ezt a vedlést számos változás előzi meg a lárva szervezetében. A kutikula alatt található összes függelék kezdete kifelé fordul, vagy kifelé mozog, és a test külső részévé válik. Ezek a szervek fokozatosan felveszik a bábra jellemző formát. A fej, a szájrészek és a lábak elülső részről hátsó hasi részre fordulnak. A gubó megpörgetése után a lárva felegyenesedik és lefagy. Megkezdődik a bábállapot előkészítési folyamata, amely jelentős időt igényel. Ezt az időszakot prepupal szakasznak nevezik.
Prepupal szakasz. A prepupa szakasz időtartama a munkásméhnél 3 nap, a királynőnél - 2, a drónnál - 4 nap. Ezt az időszakot a lárva szerkezetének jelentős átalakulása jellemzi. A lárva kutikulája teljesen elválik a bábtól egy speciális vedlési hormon, ekdizon hatására. Kivételt képeznek a lárva- és bábspirakulák közötti légcsőkapcsolatok. A fej már 2/3-a a felnőtt fejének. A szemek megnagyobbodnak, felületük begyűrődik és ráncosodik. A mellkast jól látható szűkület választja el a fejtől. A szárnyak szélességben növekedni kezdenek, ebben a szakaszban ráncosak és szorosan a mellkashoz nyomódnak. A szájrészek megnyúlnak és tömör csoportba egyesülnek.
A mellkas és a has szegmensei vízszintes vonal mentén két részre oszlanak - háti és hasi, fokozatosan elsajátítva a felnőttre jellemző szerkezetet.
Ezenkívül a belső szervek romlásnak vannak kitéve. Ezt a folyamatot hisztolízisnek nevezik. Ezt kíséri a vérsejtek - fagociták - behatolása és bejuttatása a szövetekbe. Az e folyamatok mögött meghúzódó kémiai reakciókhoz szükséges energiaforrások a lárva zsíros testében lévő zsírok, glikogén és egyéb szénhidrátok tartalékai, a hemolimfa és az izmok cukra. A hisztolízis során a hemociták felfaló sejtekként, azaz fagocitákként kezdenek működni, és az enzimek a zsírtestet, a lárva izomzatának nagy részét tápanyag-szubsztrátummá alakítják, amelyet hemolimfa juttat a növekvő szövetekbe. A hisztolízis későbbi szakaszaiban magukat a fagocitákat veszik fel az újonnan fejlődő szövetek. A legnagyobb változásokon az izomrendszer megy keresztül, aminek következtében a prepupa immobilizálódik.
Az elülső bélben billentyűs mézes golyva jelenik meg, a középső bél hurokszerűvé válik, és hajtogatott szerkezetet kap. A hátsó bélben két rész jelenik meg - a vékonybél és a végbél. A malpighi erek tubulusainak száma nő. Az idegi lánc ganglionjainak részleges fúziója figyelhető meg, melynek eredményeként a bábnak hét hasi lánc ganglionja van a lárva 11 ganglionja helyett. A kövér test mérete csökken.
A prepupa stádium a régi kutikula leválásával ér véget, és alóla kibújik a báb.
Bábszínpad. Az újonnan kikelt báb testének teljes felülete redőkkel borított, de nem ráncos. Felépítésében a báb hasonlít egy kifejlett méhhez, bár ez utóbbitól eltérően mozdulatlan, teste pigmentációtól mentes (fehér). A belső életfolyamatok azonban nem lassulnak, hanem nagy hatékonysággal haladnak. A bábot közvetlenül az utolsó vedlés befejezése után kutikula borítja. A bábállapotot mélyreható átalakulások jellemzik. Az új szervek és szövetek kialakulásához, illetve a hisztogenezishez szükséges műanyagok forrásai a hemociták által hordozott bomlástermékek. A forgó mirigyek, az emésztőcsatorna, a zsírtest és az izmok teljesen szétesnek.
A lárva elpusztult szervei és szövetei helyett egy kifejlett rovar új szervei képződnek.
A fej sokkal hosszabb, és majdnem a felnőtt fejének méretére tágult. A kitágult fej és a szűkült prothorax között híd van. Hiányzik azonban a mellkasi és a hasi szegmens között. A bábnak még nincsenek szárnyai, de az antennák, orr és lábak már jól láthatóak.
A zsírtest egyes elemei ebben az időszakban a fejben és a mellkasban szétszóródnak, egy részük pedig a hasban koncentrálódik. Az életkor előrehaladtával a zsírtest elemei egyenletesebben oszlanak el a fejben, a mellkasban és a hasban. A gyomor szerkezete megváltozik.
A szív alig változik: a kamrák száma 12-ről ötre csökken. A bábállapotban a szív nem hagyja abba a működését, mivel a metamorfózis folyamatához a hemolimfa mozgása szükséges. A fejben lévő supraesophagealis ganglion tömege megnő. Ez a differenciálatlan idegsejtek miatt következik be. Ezenkívül a mellkasban lévő ideg ganglionok egyesülnek, valamint az utolsó négy ganglion a has végén.
Az ivarmirigyek kezdetlegessége nem pusztul el. Növekedésük kezdeti szakaszában a zsírtest sejtjeinek szétesésével és a műanyagok albumidok formájában történő felszabadulásával jár. Fejlődésük során érintkezésbe kerülnek az ektodermából kiinduló ivarmirigycsatornák részeivel.
A chrysalis belső életének egyik mutatója a légzése.
A légzőrendszer számos változáson megy keresztül a hisztolízis és a hisztogenezis során. Tehát az első és a harmadik mellkasi spirálok nőnek, a második pedig éppen ellenkezőleg, csökken. Elpusztult, légcső. Képzeletbeli primordiumokból új légcsövek, valamint légzsákok keletkeznek, amelyek a légcsőtörzseken helyezkednek el. 1
Az összetett szemek a fej oldalán lévő rügyekből nőnek. Mint fentebb említettük, a báb kezdetben fehér. A fejlődés során a különböző szervek képződése során a báb külső héjának színe a következő sorrendben változik: a bábozás után egy nappal az összetett szemek még tiszta fehérek, 2 nap múlva sárgás árnyalat jelenik meg a bőrön. A 14. napon rózsaszínűvé válnak, a 16. napon pedig sötétlilává.
A lábak ízületei eleinte sárgásak, a 18. naptól a lábakon lévő ízületek és karmok sárgásbarna színt kapnak, a sarok és a szájrész valamivel világosabb. A mellkas elefántcsontszínűvé válik. A 19. naptól a mellkas enyhén sötétedni kezd. Hasa elefántcsont. A lábak szegmenseinek végei barnák. Jelentősebb sötétedés figyelhető meg az áll végén és a felső állkapocs tövében. A szemek lilává válnak. A 20. napon az egész báb színe sötétszürke.
Imago színpad. A bábállapot végén lehullik a bábbőr, amely alól egy teljesen megformált méh bújik elő. A méh az állkapcsával átrágja a sejt fedelét, és a lép felületére emelkedik. Az újonnan kikelt méh egy idősebb méhhez képest puhább kitines borítású, ráadásul testét sűrűn borítják szőrszálak. Idővel a méhek elveszítik hajszáluk egy részét, és a kitin sokkal keményebbé válik. A képzeletbeli időszakban a méhegyedek külső szerkezetében nem történik jelentős változás. "
Anyagcsere a posztembrionális időszakban. A lárva és a báb intenzív anyagcsere-folyamatokon megy keresztül, melynek eredményeként felhalmozódnak a felnőtt méh kialakulásához szükséges tápanyagok.
Strauss (1911) szerint a méhek anyagcseréjének sajátossága a jelentős mennyiségű glikogén felhalmozódása a metamorfózishoz szükséges energiaként. A lárvaállapot végére a glikogéntartalom eléri a lárva száraz tömegének 30%-át. Ezzel párhuzamosan a zsír mennyiségének növekedése, a zsírsejtek növekedése.
Először is, a glükózt a méhek szervezete fogyasztja. Összes tartalma a munkásméh-lárvák szervezetében kétszer akkora, mint az anyalárvák szervezetében. Amikor a hemolimfában a glükóztartalom a megengedett érték alá esik, az összetett szénhidrátok elkezdenek lebomlani és felhasználódnak. A glükóz legmagasabb koncentrációja a nyitott szakasz közepén figyelhető meg.
Megjegyzendő, hogy a legtöbb vizsgált enzim aktivitása a fiatal lárvákban a legmagasabb. Ezután csökken, és a bábállapotban eléri a minimumot. Ugyanakkor a munkásméhekben az enzimaktivitás fejlődése jelentősebb, mint a királynőké. Egyes vélemények szerint a posztembrionális fejlődés időszakában a királynők szénhidrát-anyagcseréje aerobabb a dolgozó egyedekhez képest, és a királynők felgyorsult fejlődése számos enzim magasabb aktivitásával magyarázható. A mitokondriumok (sejtszervecskék) mennyiségi és minőségi különbségei a nál idősebb méhlárvákban
- naposak az azonos korú munkásméh-lárvákéhoz képest. Ennek megfelelően a mitokondriumokban lezajló egyes oxidatív folyamatokban is megfigyelhető némi eltérés.
A királylárvában az oxigénfogyasztási görbe felfelé megy, amíg a sejt le nem záródik. A lezárást követően azonban az oxigénfelvételi görbe leesik (ami egybeesik a gubópörgéssel), és a kezdeti bábstádiumban eléri a minimumot. Az egységnyi tömegre jutó oxigénfogyasztás szintje a királynőknél a legalacsonyabb a dolgozó méhekhez és drónokhoz képest.
Az egyedenként felszívódó oxigén mennyisége a drónokban a posztembrionális időszakban magasabb, mint a királynőké és a munkásméhekben.
A légzési együttható, amely a munkásméhben a lárva teljes fejlődési szakaszában az energiaforrásként használt szubsztrátot jellemzi, változik, értéke meghaladja az egyet. Ha a légzési együttható értéke egynél nagyobb, akkor ez az oxigénben gazdag vegyületek oxigénben szegényebb vegyületekké való átalakulásával kapcsolatos folyamatok intenzitását jelzi.
A légzési együttható növekedése a lárvafejlődés második periódusában egybeesik a minőségileg eltérő típusú táplálékra, főként szénhidrátra való átállás idejével. A sejt lezárása után és a lárvák bebábozódásáig a légzési együtthatót 0,9-re állítjuk be.
A lezárt lárvában és prepupa fő energiaforrásai a glikogén formájában lerakódott szénhidrátok, a bábállapot második felében pedig a zsírok.
A méhben a munkásméhtől eltérően a lárvafejlődés 2. napjára csökken a légzési együttható, ami a zsíranyagcserére jellemző. Ugyanezen stádium 4-5. napján a légzési együttható közel megegyezik a munkásméh-lárvák légzési együtthatójának értékével, ami a szénhidrátok zsírrá alakulását jelzi. A zárt lárva állapotában a légzési hányados egy alatt van, ami a szénhidrátok lebomlását jelzi.
A fentiekből arra a következtetésre juthatunk, hogy a mézelő méhekben a gázcserét számos, a rovarokban rejlő mintázat jellemzi. A lárvák fejlődése során a légzés intenzitása, amelyet az egységnyi tömegre jutó oxigén mennyiségében fejezünk ki, csökken. A lárvánként elfogyasztott oxigén mennyisége azonban növekszik, és a bábozás előtt eléri a maximumot. A bábállapotban a gázcsere csökken, a felnőtt kilépése felé fokozódik. Ugyanakkor a hasonlóságok mellett különbségek mutatkoznak a mézelő méhek három formája légúti cseréjében.
A rovaroknál a fejlődés két időszakra oszlik - embrionális és posztembrionális.
Embrionális fejlődés. A rovarpetesejtek többnyire ovális alakúak, és nagyon gazdag sárgájában, amely kitölti a tojás teljes központi részét. Ennek megfelelően a tojás zúzása részleges, felületes; ennek eredményeként azonos sejtekből álló folyamatos réteg, a blastoderma képződik, amely a központi sárgáját borítja. Ugyanakkor a magok egy része a tojássárgája tömegében marad: a jövőben speciális tojássárgája sejtek képződnek ezek költségén. Az embrió leendő ventrális oldalán a blastoderma megvastagszik. Ezt a területet csíracsíknak nevezik; a jövőben ennek költségén kialakul az embrió testének fő része.
A csíracsík régiójában megindulnak a differenciálódási folyamatok. A szaporodás és a blastoderma sejtek mélyére való behatolás eredményeként két sejtcsík képződik - mezodermális csíkok, és maga a blastoderma válik ektodermává. Később a mezodermális csíkok páros primordiumokká osztódnak, így kettős coelomikus zsáksor keletkezik. Magasabb rovaroknál (pl. Kétszárnyúak) a mezodermális csíkok szegmentációja elnyomódik, és a független coelomok szétválása nem következik be.
A rovarok endoderma kialakulásának kérdése még nem teljesen megoldott. A vitellin sejtek számos formában az endoderma kezdetleges elemének tűnnek. Nál nél Kétszárnyúakés néhány más rovar esetében két sejtcsoport különül el a csíracsík elülső és hátsó végén, amelyeket az endoderma kezdetének is tekintenek.
Már a csírarétegek lerakásának legelején megkezdődik az embrionális membránok kialakulásának folyamata, amely rendkívül jellemző a rovarok fejlődésére.
A legegyszerűbb és legelterjedtebb esetben a csíracsík perifériája mentén az ektoderma először egy görgőt, majd egy kétrétegű redőt alkot, amely minden oldalról fokozatosan a csíracsíkig növekszik, amely egyúttal némileg belemerül a csíracsíkba. a sárgáját. Végül a redő szélei az embrió rudimentuma fölé záródnak, és maguk és közötte marad a folyadék által elfoglalt magzatüreg, amelyet kettős membrán választ el a külső környezettől. A membrán külső rétege az embrió fölé záródó ektodermális redő külső rétegeinek összeolvadása eredményeként jön létre, és serosa-nak nevezik, a belső réteget ugyanannak a redőnek a belső rétegéből, ez utóbbi közvetlenül korlátozza a magzatüreget. és amnionnak hívják. A serosa és az amnion megvédi az embriót a károsodástól.
Eközben a csíracsík szegmentálódik, és a szegmensek elszigeteltek, mint a többiben Ízeltlábúakés gyűrűk, sorrendben elölről hátrafelé. A fejrészben a szemészeti és antennalebenyeket a megfelelő rudimentekkel, az interkaláris szegmenst, amely ezt követően redukálódik, és a szájvégtagok 3 szegmensét helyezik el. Tovább hátul 3 mellkasi, a végtagok alapjait hordozó és 11 hasi szegmens különbözik.
Ami a belső szervek differenciálódását illeti, főbb jellemzői a következők. Az elő- és a hátsó bél két ektodermális invaginációval jön létre. A középbél eredetének kérdése bonyolultabb. A rovarokat az utóbbi kialakulásának elképesztő sokféle módja és forrása jellemzi. Egyes esetekben a tojássárgája endodermális sejtjei miatt képződik, másokban az ektodermális elemek vesznek részt a középbél kialakulásában - speciális sejtek, amelyek az elülső és a hátsó bél rudimentumainak vakon zárt végein lokalizálódnak. És végül számos fajnál (parthenogenetikus nőstény levéltetvek, egyes ichneumonok) a középbél csak az ektoderma révén alakul ki, az előbél anlage erős növekedése miatt, amely eléri a test hátsó végét. Az endodermális elemek ugyanakkor degenerálódnak. A malpighi erek a hátsó bél ektodermájából származnak. Az idegrendszer egy ektoderma hasi gerince formájában helyezkedik el, amely azután az integument alá süllyed. A cölomikus zacskók szétesnek, vegyes testüreget - mixocoelt - képeznek. Az egyidejűleg felszabaduló sejtelemek izmokat, szívet, zsírtestet és néhány más szervet eredményeznek. Az elsődleges csírasejtek nagyon korán rakódnak le, néha még abban az időszakban is, amikor a sárgáját egyetlen réteg blastoderma veszi körül.
Az embrió növekszik, a tápláló sárgáját folyamatosan fogyasztják, és az embrionális membránok szétrepednek, a hátoldalon redők formájában beburkolódnak, amelyek degenerálódnak. Az embrió készen áll a tojásból való kiemelkedésre, és ezzel véget ér a rovar embrionális fejlődése.
Posztembrionális fejlődés. A rovaroknál egy fiatal állat vagy egy lárva, teljes számú szegmenssel emelkedik ki a tojásból. Az egyetlen kivétel a primitív cryptomaxillárisok, amelyek a neg-hez tartoznak. Protura, amelyben a fiatalkorúak három hátsó hasi szegmens hiányában különböznek a felnőttektől. Ez utóbbiak a hátsó növekedési zónából alakulnak ki egymás után az első vedlés után. Így a rovarokra jellemző az anamorfózis nélküli fejlődés, és csak a proturban található a maradék anamorfózis.
A rovarok posztembrionális fejlődését jelentős sokféleség jellemzi, amelyet nagymértékben meghatároznak ezen állatcsoport képviselőinek biológiai jellemzői.
A legprimitívebb formákban (subcl. Entognatha) egy fiatal szervezet növekedését és fejlődését nem kísérik jelentős szerkezeti változások. A fiatal egyedek minden fő jellemzőjükben hasonlóak az anya szervezetéhez, azaz hiányzik a lárvaállapot és a metamorfózis. Lényegében ebben az esetben közvetlen fejlesztésről van szó. A szárnyas rovarok, amelyek evolúciós szempontból magasabbak, éppen ellenkezőleg, metamorfózissal rendelkeznek, és az átalakulás természete szerint félmetabolikusra - hiányos átalakulással és holometabolikusra - teljes metamorfózissal oszthatók. A tökéletlen metamorfózis a szárnyas rovarok alacsonyabban szervezett csoportjaira jellemző: orthoptera, csótány, poloska, májusi, szitakötő. A tojásból kikelő lárva tökéletlen átalakulás esetén általános megjelenésében már sok tekintetben hasonlít a kifejlett szervezethez, főként a szárnyak embrionális állapotában, a másodlagos ivartulajdonságok fejletlenségében, és gyakran valamilyen átmeneti állomány jelenlétében tér el tőle. , tisztán lárvaszervek: például légcső-kopoltyú a májusi légkör lárváinál stb. A szárnyak eleinte csak alig látható külső rudimentek formájában vannak. A lárva és a kifejlett szervezet közötti különbség kiegyenlítése fokozatosan történik, számos vedlés kíséretében.
Minden egyes következő vedléssel a lárva egyre jobban hasonlít a rovarok kifejlett állapotához, vagy imágóhoz, megnő a szárnyak kezdetlegessége stb.
Egyes primitív rovaroknál a vedlések száma nagyon nagy, és elérheti a 25-30-at is (május legyek). A többi különítmény képviselőinek kevesebb sora van (4-5).

A holometabolikus rovaroknál a fejlődés eltérő. A lárva élesen eltér a kifejlett állattól: egyáltalán nincs külső szárnya, gyakran eltérő számú végtaggal rendelkezik stb. A lárva karakterei a vedléssorozat során, azaz a lárva teljes lárvakorszaka alatt megmaradnak. élet. A holometabolikus rovarok lárvái rendkívül változatosak. A lárva számos primitív tulajdonság meglétében különbözik az imágótól: a boncolás nagyobb homonómiája, a szárnyak hiánya, az érzékszervek gyenge fejlettsége (nincs összetett szem, csak szemhéjak, az antennák nagyon kicsik, stb.), és sok esetben a végtagok eszközét és számát is. Nagyon gyakran a lárvakorban a száj végtagjai egy típus szerint vannak elrendezve (lepke hernyók rágása), felnőtteknél eltérően (felnőtt pillangók szopása).
Mint már említettük, a lárvára jellemző szerkezet a lárva teljes periódusa alatt megmarad: az átalakulás nem fokozatosan megy végbe, hanem a metamorfózishoz szükséges változások a lárva utolsó vedlési idejére tolódnak el. Ebben az időszakban a lárva szervezetében olyan alapvető átalakulások mennek végbe, hogy a megszokott élettani funkciók, különösen a táplálkozás és a mozgás folytatása megnehezül, sőt lehetetlenné válik. Ez abban nyilvánul meg, hogy a lárva az utolsó vedlés után nyugalmi állapotba kerül - krizálissá válik. A báb a holometabolikus átalakulás legjellemzőbb szakasza. Minél teljesebb a nyugalmi állapot, annál jobban kifejeződik a teljes átalakulás.
Egy rovar egyedfejlődése vagy ontogenezise 2 periódusra osztható - a tojáson belüli vagy embrionális fejlődésre és a tojás elhagyása utáni vagy posztembrionális fejlődésre.
A legtöbb rovar tojást rak. A rovartojás olyan sejt, amely magot, protoplazmát, deutoplazmát vagy sárgáját tartalmaz, amely szükséges az embrió táplálkozásához és fejlődéséhez. Ezenkívül a rovarpeték gyakran tartalmaznak szimbiotikus mikroorganizmusokat, amelyeket az anyától a petefészken keresztül (azaz transzovariálisan) nyernek. A tojást kívülről chorion - héj borítja. A chorion néha különálló mikroplasztikával rendelkezik, amely megbízható jelként szolgál a rovarnemzetségek, sőt -fajok megkülönböztetésére a tojás fázisa szerint. A chorion alatt található a tojás valódi vagy sárgája membránja. A chorion kialakulásával egy nyílást hagynak a mikropilának, amely a spermiumok átjutását szolgálja a megtermékenyítés során. A tojás elülső és hátsó végei megfelelnek az embrió helyzetének. A petefészekben a petesejt elülső pólusa az anya feje felé irányul, illetve a pete dorsalis és ventrális oldala található.
A tojások mérete és megjelenése változatos. A méret levéltetveknél és tripszeknél 0,02-0,03 mm, sáskánál 8-10 mm. A tojás levéltetvekben sima, gombócban bordázott, fehérje, néha fedővel ellátva, ráncokkal borított.
Tojás alakú. Ovális - bogarak, pillangók - hosszúkás, félgömb alakú - gombóc, palack alakú - nappali pillangók, hordó alakú - poloskák, száruk van - fűzős, balekos. Petetartás: egyenként, csoportosan, nyíltan vagy szubsztrátumban, védett, ivarmirigyváladékkal borított.
Az embrionális fejlődés a sejtmag feldarabolásával és az így létrejövő leánymagok kis protoplazmaterületekkel történő mozgásával kezdődik a tojás perifériájára. Itt a leánymagok tömegéből egy folyamatos sejtréteg képződik - a blastoderma. Ezt követően a blastoderma csíra- és extraembrionális zónákra differenciálódik: az utóbbi sejtjei nem vesznek részt az embrió kialakulásában, míg a csírazóna sejtjei intenzívebben kezdenek osztódni, és embrionális sávot alkotnak a ventrális oldalon. a tojásból, amely azután kinyúlik, és ektoderma és mezoderma keletkezik.
Az embrió fejlődését blasztokinézis kíséri: embrionális membránok képződnek, a fej, a mellkasi és a hasi szakaszok elkülönülnek. A belső szerkezet alapjai a mezoderma kialakulásával kezdődnek. Az ektodermából származik az összes külső burkolat - majd a száj és a végbélnyílás, az elülső és a hátsó belek. A mezoderma az izomrendszert, a zsírtestet, a háti ereket, a nemi mirigyek hüvelyét eredményezi.
Az embrió mozogni kezd, levegőt, folyadékot fog fel a tojásból, a chorion eltörik, és a lárvává változott embrió kijön.
2. A rovarok szaporodása számos olyan tulajdonsággal rendelkezik, amelyek biológiájuk lényeges részét képezik, és megfontolásukat igénylik. Ezek a jellemzők magukban foglalják a szaporodási módszereket, a kiegészítő táplálkozást, a nemek találkozását és a megtermékenyítést, a szexuális termelékenységet.
Szaporodási módszerek .
A legtöbb rovarnál a szaporodás párosodással és megtermékenyítéssel jár, azaz. két nem részvételével kapcsolatos - ezért biszexuális (biszexuális) gamogenetikusnak nevezik. A legtöbb rovar tojást rak, de vannak más szaporodási módszerek is: élve születés, partenogenezis, pedogenezis, poliembriónia. Az élve születés azt jelenti, hogy az embrionális fejlődés az anya testében teljesedik ki, ezért a tojásrakás helyett lárvák (levéltetvek, légy, vérszívó legyek) születnek a világra. A lárvák a tojáscsövekben vagy a petevezetéken való áthaladás közben kelnek ki.
Parthenogenezis - a szűz szaporodást a megtermékenyítés hiánya jellemzi, és mind a petesejt, mind az elevenszülő rovaroknál megfigyelhető.
A partenogenezis formái:
1. arrhenotokia- megtermékenyítetlen petékből csak a hímek fejlődnek ki; 2) thelytokia - megtermékenyítetlen petékből csak a nőstények fejlődnek ki; 3) amphitoky- a hímek és a nőstények megtermékenyítetlen petékből fejlődnek.
A partenogenezis lehet fakultatív, állandó és ciklikus.
Fakultatív partenogenezis vagy spadic inkonzisztensen jelentkezik, külső hatásokra vagy a tojásrakó nőstény bizonyos fiziológiás állapotánál jelentkezik (cigánylepke, nyárlepke).
Az arrhenotoky típusú állandó partenogenezis jellemző a társas rovarokra. A méhekben a megtermékenyítetlen petékből mindig hím, a megtermékenyített petékből pedig nőstények fejlődnek. A thelytoky típusú permanens partenogenezis olyan rovaroknál figyelhető meg, amelyek nem rendelkeznek koloniális életmóddal. Így sok féregben, tripszben, fűrészlegyben és néhány lovasban hiányoznak a hímek, a szaporodás mindig partenogenezis útján történik, és csak a nőstények kelnek ki a megtermékenyítetlen petékből.
A partenogenezis nagyon fontos szerepet játszik a rovarok életében. A partenogenezisnek és annak módosulásainak köszönhetően a faj szaporodási potenciálja ugrásszerűen megnő: partenogenetikus szaporodással jelentősen megnő a faj túlélési esélye kedvezőtlen körülmények között és a megtelepedés során.
3. A tojás elhagyása után megkezdődik a rovar posztembrionális fejlődése. Ez az időszak nem egyszerű növekedés és testméret növekedés, hanem a test egyik fázisból a másikba való átmenete jellemzi. Ezt a fajta egyedfejlődést ún metamorfózis vagy fejlődés átalakulással.
A metamorfózis lényege abban rejlik, hogy a fejlődő egyed élete során jelentős átalakuláson megy keresztül morfológiai szervezetében és biológiájában. Ebben a tekintetben a posztembrionális fejlődés 2 fázisra - lárva és kifejlett - differenciálódik. A lárva fázisban az egyedek növekedése és fejlődése, az imágó fázisban a szaporodás és a megtelepedés következik be. Más esetekben e fázisok között egy köztes fázis következik be - a báb. A metamorfózisnak 2 fő típusa van - a hiányos és a teljes átalakulás. hiányos átalakítás ill hemimetamorfózis 3 fázis - tojás, lárva, kifejlett. A lárvák az imágóhoz hasonlóak (összetett szemek, szájrészek, szárnyak alapjai, hasonló életmód), imágószerűek. Teljes (holometamorfózissal) 4 fázis. A lárvák egyáltalán nem hasonlítanak a felnőttekre, nincsenek összetett szemek, szárnyak, különböző körülmények között élnek.
Némelyiknek hasi lába van, más a szája, másoknak selyem- vagy pókmirigyük van.
A hiányos átalakulás módosulásai: a) hipomorfózis - leegyszerűsített hiányos átalakulás, amely a szárnyakat vesztett szárnyas rovarokra jellemző (másodlagos szárnyatlan tetvek, sáskák, szöcskék, tücskök, csótányok, pálcikaevők, csótányok); b) hipermorfózis - bonyolult, nem teljes ciklus (aleurodidok, tripszek, kokcid hímek). A nyugalmi állapot lárváinak megjelenése a fázis végén.
Anamorfózis- a lárvák hasonlóak a kifejlettekhez, de kisebb számú hasi szegmensük van, a lárva fejlődésével további szegmensek nőnek a has felső részén, de a teljes szám csak a kifejlett fázisban éri el.
protomorfózis– vedlés kifejlett állapotban, a lárva némi hasonlósága a kifejlett fázishoz, de a lárva mellkasra és hasra (podura, kétfarkú, sörtefarkú) nem osztódik.
hipermetamorfózis- a teljes átalakulás komplikációja (2 lárvaforma jelenléte, és néha bábok (hólyagok)).
4. A lárva fázis közvetlenül a tojásból való kikelés után következik be. Kezdetben világos, kikelés után a lárva színtelen, puha borítású. A nyíltan élő rovaroknál a lárva gyorsan elsötétül és megkeményedik. Az embrionális tojássárgája maradványainak emésztése és az ürülék kiürítése után táplálkozási ingerek jönnek létre. A lárva egy pár fokozott táplálkozás, növekedés és fejlődés. Növekedésüket és fejlődésüket időszakos vedlés, a bőr kutikula leválása kíséri – a vedlés miatt a test megnövekszik és jelentős változások következnek be. A linkek száma változó: 3 (legyek), 4-5 (bogár), 25-30 (padenki).
Minden vedlés után a lárva a következő stádiumba, vagyis korba lép, ezért a vedlések megosztják a lárvák korát. A vedlésszámnak megfelelően a lárvaállatok száma is megtalálható. A vedlések korának meghatározása (amely a küzdelem időpontjának megtervezéséhez szükséges) a következő jellemzők szerint történik: antennák a sáskáknál, testméret növekedés (a fejkapszula szélessége) pillangó hernyóknál és bogárlárváknál . A lárvák 2 típusúak - felnőtt (elsődleges) és másodlagos.
Naiads - kopoltyús lárvák, vízben élnek (szitakötők, padyonki, kőlegyek).
Campodeoid - mozgékony, sötét színű, sűrű béléssel és 3 pár mellkasi lábbal, jól izolált prognatikus fejjel (földi bogarak, úszók, fűzők).
Féregszerű - ülő, világos színű, hasi lábak nélkül.
A hernyóknak jól elkülönülő fejbőrük van, 3 pár mellkasi, 2-8 pár hasi.
bábfázis. A metamorfózisnak ez a fejlődési szakasza csak a teljes ciklusú rovarokra jellemző. A báb jellemzője, hogy nem tud táplálkozni, és nagyon gyakran mozdulatlan marad. A lárva által felhalmozott tartalékokból él, és gyakran alvó fázisnak tekintik.
Külsőleg a báb nem úgy néz ki, mint egy felnőtt, de számos jele van a felnőtt kornak: szárnyak, lábak, antennák külső elemei, összetett szemek stb. A hisztolízis és hisztogenezis a bábfázisban történik.
Hisztolízis- a lárva belső szerveinek szétesése, amelyet a vérsejtek - hemociták - behatolása és szöveteibe való behatolása kísér. A hemociták sejtfalóként működnek, azaz. fagociták, amelyek fokozott aktivitása a szövetanyag pusztulásához és felszívódásához vezet. A hisztolízis megfogja az izomrendszert, így a prepupa mozdulatlanná válik, hatással van az emésztőrendszerre, de nem érinti az ideg- és reproduktív rendszert, valamint a háti eret.
Hisztogenezis- a képzeletbeli élet szövetei és szervei létrehozásának folyamata. Ezen új szövetek és szervek kialakulásának forrásai a hisztolízis termékei. A hisztogenezis átfogja az izom- és emésztőrendszert, átrendezi őket új, képzeletbeli funkciókra. Az izomrendszer újjáépül a repülés új mozgásformáihoz, az emésztőrendszer pedig az új típusú táplálékokhoz.
A hisztogenezis során a fő szerepet a képzeletbeli rudimentumok játsszák - hipodermális sejtcsoportok, amelyekből bizonyos szövetek és szervek származnak.
A vedlési hormon az exidon, amely a lárvák normális fejlődéséhez szükséges.
Vannak a következők báb típusok:
nyitott bábok- szabad, csak a testhez nyomott képzeletbeli függelékek (antennák, lábak, szárnyak), jellemző a bogarakra.
Mozgatható állcsontú bábok – mozgatható felső állkapcsokat használnak a gubóból való kilépéshez, és maguk is tudnak mozogni (hálós, körömlepkék, fogazott lepkék). A mozdulatlan állcsontú bábok a gubóból kibújva ez utóbbit nem tudják felhasználni (bogarak, hártyásszárnyúak, nagyszárnyúak, sok kétszárnyú).
Rejtett babák edzett, nem hulló lárvabőr borítja, amely héj vagy hamis gubó szerepét tölti be - puparia (legyek).
fedett- képzeletbeli függelékei szorosan a testhez vannak nyomva és forrasztva, mivel az utolsó vedlés során a lárva olyan titkot választ ki, amely megszilárdulva kemény héjjal borítja be a bábot (pillangók, katicabogarak).
A bábozás előtt egyes lárvák gubóval (selyem, pókháló) veszik körül magukat. Néha a bábozás helye növényi szár, földbölcső, nyitott bábozás.
Imago fázis.
A bábból kibújó rovarokon felnőtt jelei vannak, de eleinte a szárnyak össze vannak hajtva. Aztán széttárják szárnyaikat, a bőrszövet megvastagszik, és olyan színt kap, mint egy teljesen kifejlett kifejlett rovar. A kifejlett fázisban a rovarok nem vedlődnek és nem képesek növekedni. A kivétel a padyonki, a bolondok. A felnőtt fázis biológiai funkciója a szétszóródás és a szaporodás. Ez már a faj életének függvénye, és a faj létének fenntartását célozza. A szárnyaknak köszönhetően a felnőtt rovarok letelepedési képessége jelentősen megnő, és a szaporodás lehetővé teszi, hogy a leszármazottakat új helyen hagyják.
A kifejlett rovarok elterjedése aktív és passzív repüléssel egyaránt megtörténik. Az aktív repülés általában a nagy rovarokra jellemző, és számos szitakötő-, sáska-, pillangó-, bogara-fajnál megfigyelhető, és széles körben elterjedt. A passzív repülés a levéltetvekre és legyekre jellemző.
A felnőttekbe való átmenetet a testszín külső változásai, a nőstény has méretének növekedése kíséri, petével teli petefészkek fejlődése miatt - termeszek és hangyák megtermékenyített nőstényeiben - szárnyak leesése, éretlen sáska - rózsaszín , érett - élénksárga.
A hímek és a nőstények szexuális dimorfizmusa számos külső, másodlagos nemi jellemzőben - az antennák alakjában és méretében, a testméretben, a különböző szerkezeti részletekben (orrszarvúbogár, szarvasbogár) - különbözik. A hímek mozgékonyabbak, nyitottabb életmódot folytatnak.
Polimorfizmus - a rovarok létezése, ugyanazon faj külső formái (hangyák, méhek, termeszek, hímek, nőstények, munkások, katonák). A szexuális polimorfizmus a családon belül szabályozott, és nem függ külső tényezők hatásától.
Az ökológiai polimorfizmus a külső környezet (hosszúszárnyú, rövidszárnyú, szárnyatlan poloskák) hatására jön létre.