Embrionalni razvoj žuželk se običajno začne s površinskim drobljenjem jajčeca, kar je povezano z bogato vsebnostjo zadnjega hranljivega rumenjaka. Bastoderma, ki nastane na površini jajčeca, je na nekem delu površine debelejša; ta odebelitev povzroči nastanek zarodne črte in opredeljuje trebušno stran zarodka.
Obročasta guba, ki se oblikuje ob robu zarodnega traku, prerašča in raste skupaj nad zarodnim trakom, daje dve plasti zarodnih membran - notranjo (amnion) in zunanjo (serozo). Iz celične plasti zarodnega traku na strani, obrnjeni proti rumenjaku,primarni endoderm se loči - skupni zametek endoderma in mezoderma. Kasneje se zarodna črta segmentira.
Postembrionalni razvoj žuželk je zelo raznolik zaradi različnih pogojev za obstoj mladic žuželk in stopnje njihove prilagoditve nanje. Razlikujemo lahko dve glavni vrsti postembrionalnega razvoja: nepopolno transformacijo, hemimetabolizem, in popolno transformacijo, holometabolizem. Za holometabolizem so značilne globoke spremembe pri mladičih, zaradi česar se mladiči po strukturi močno razlikujejo od odrasle žuželke (odrasle osebe). Zato prehod iz stanja mladičev, to je iz larvalne faze v imaginalno fazo, zahteva korenito prestrukturiranje organov; to se doseže s sodelovanjem vmesne faze med fazo ličinke in imaginalno fazo, med katero ostanejo mladiči v stanju mirovanja; ta vmesna faza je lutka. Med pupalnim stanjem se notranji organi žuželke bolj ali manj prestrukturirajo ali pa se popolnoma uničijo (histoliza) in se ponovno zgradijo; v slednjem primeru pride do nastajanja organa na novo zaradi tako imenovanih imaginalnih diskov, sestavljenih iz celic embrionalne narave (histoblastov). Tako je pupalno obdobje obdobje aktivne histolize in histogeneze (tkivne neoplazme). Pupa navzven predstavlja imaginarno fazo z značilnimi organi, ki pa so ostali premalo razviti.
Razvoj v višjih razredih žuželk je holometaboličen pri hroščih, lepidopterah, tulcih, bolhah, himenopterah in dvokrilcih.
V primeru nepopolne preobrazbe - hemimetabolizma - ni tako globoke razlike v zgradbi mlade in odrasle faze: juvenilna je zdaj bolj, nato nekoliko manj po zgradbi podobna imaginarni fazi.Nazadnje, v primerih, ko prehod iz mladega faza do odrasle osebe se zmanjša le na rast in izločanje (v povezavi z rastjo) starega pokrova (exuvia), govorijo o amebolizmu, to je odsotnosti preobrazbe; primer amebolizma je razvoj Collembola med Apterygota. Običajno pa pri zaporednih stadijih, ki so med seboj ločeni z linjanjem, to je z odpadanjem pokrova prejšnje starosti, pride tudi do sprememb v obliki organov: na primer dolžina anten in cerkov relativno poveča, število segmentov se poveča; nekateri organi se lahko ponovno pojavijo, na primer zunanji spolovili. Takšen je razvoj pri nekaterih Apterygota, kot sta Thysanura in Diplura, in pri večini hemimetabolnih Pterygota. Pri nekaterih Pterygotah, ki se razvijajo s hemimetabolizmom (kačji pastirji, enodnevnice; kamenjarji in deli členonožcev), je razvoj zapleten zaradi dejstva, da mladiči živijo v drugačnih razmerah kot odrasli; tako se v vodnih telesih razvijejo mladiči kačjih pastirjev, enodnevnic in kamenčkov, ki v zvezi s tem dobijo organe za vodno dihanje - škrge, ki so primer ličink (provizornih) organov. Pogoji obstoja se odražajo tudi v razvoju drugih organov ličinke, ki se prilagajajo okolju, na primer preoblikovanje sprednjih nog mladoletne cikade v organ za kopanje v povezavi z njenim podzemnim življenjem.
Razvoj žuželk
Individualni razvoj žuželk (ontogeneza) je sestavljen iz embrionalnega razvoja, ki poteka v fazi jajčeca, in postembrionalnega razvoja - po tem, ko ličinka zapusti jajčece, dokler ne doseže odrasle faze - imago.
Embrionalni razvoj. Jajčeca žuželk se razlikujejo po obliki zaradi prilagoditev na okolje, v katerem se razvijajo. Na primer, pri hroščih so jajčeca pretežno ovalna in se pogosteje razvijajo v zaprtem substratu; v stenicah - v obliki soda, pritrjen na podlago; pri metuljih - v obliki kupole ali v obliki steklenice; pri čipkah (zlate oči) jajčeca s pecljem. Jajca pogosto odlagajo v skupinah. Jajčne sklopke so lahko odprte ali zaprte. Primer odprtega odlaganja so jajčeca koloradskega hrošča, ki jih samica prilepi na spodnjo stran krompirjevih listov. Zaprte sklopke vključujejo jajčne stroke akridoidov, ki nastanejo iz delcev prsti, cementiranih z izločki adneksalnih žlez samic. Ščurki odlagajo jajca v ooteke – jajčne kapsule, ki nastanejo v ženskem genitalnem traktu.
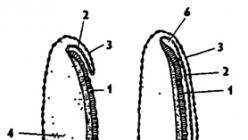
Jajčeca žuželk so zunaj pokrita z lupino - horionom, ki jih varuje pred izsušitvijo (slika 337). Na površini lupine je mikropil - majhna luknja s kompleksnim "čepom" s tubulom v notranjosti za prodiranje semenčic med oploditvijo. Pod horionom je tanka rumenjakova membrana, pod njo pa gosta plast citoplazme. Osrednji del citoplazme je zapolnjen z rumenjakom. Citoplazma vsebuje jedro in polarna telesca.
Drobljenje je površinsko. Sprva se jedro večkrat deli, hčerinska jedra z deli citoplazme migrirajo na obrobje jajčeca, so prekrita z membrano in nastane površinska plast celic - blastoderma, rumenjak pa ostane v središču jajčeca. jajce. Na ventralni površini blastoderma so celice višje in tvorijo zgostitev - zarodni trak. Ta stopnja embrionalnega razvoja žuželk ustreza blastuli.
Delitev celic zarodnega traku vodi do razvoja zarodka. Zarodni trak postopoma potone in tvori ventralni žleb.
riž. 337. Zgradba jajčeca žuželke (iz Bei-Bienka): 1 - mikropil, 2 - horion, 3 - rumenjakova membrana, 4 - jedro, 5 - polarna telesa, 6 - rumenjak

Gube blastoderma nad brazdo se zaprejo in nastanejo zarodne membrane: seroza in amnion (slika 338). Tu se kaže konvergenca z višjimi vretenčarji, ki imajo prav tako podobne lupine. Zaradi nastale amnijske votline je zarodek znotraj jajčeca v suspendiranem stanju, kar ga zanesljivo ščiti pred mehanskimi poškodbami.Poleg tega tekočina, ki napolnjuje amnijsko votlino, olajša presnovne procese zarodka.
Zarodni trak se nadalje loči v dve plasti: spodnjo - ektoderm in zgornjo - endomezoderm. Entomezoderm pri različnih vrstah žuželk lahko nastane na različne načine: z invaginacijo ali imigracijo celic.
Na naslednji stopnji razvoja se ektodermalna plast traku začne upogniti navzgor ob straneh, nato pa se zapre na zadnji strani in tvori zaprto steno zarodka. Ko se stene telesa na hrbtu zaprejo, del rumenjaka in rumenjakovih celic vstopi v telo zarodka. Hkrati s tvorbo sten telesa zarodka se v entomezodermu ločita dve skupini celic na sprednjem in zadnjem koncu telesa. To sta dva rudimenta srednjega črevesa. Nato se iz teh dveh primordijev začneta oblikovati sprednji in zadnji del srednjega črevesa, ki se nato združita. Istočasno se na sprednjem in zadnjem koncu telesa zarodka oblikujejo globoke invaginacije ektoderma, iz katerih nastanejo sprednji in zadnji del črevesja. Nato so vsi trije deli povezani in tvorijo črevesno cev.
Mezodermalni trak razpade na parne metamerne zametke celomskih vrečk. Kasneje pa razpadejo in iz mezoderma nastanejo mišice zarodka, somatska plast celomičnega epitelija, srce, maščobno telo in spolne žleze. Visceralna plast kolomičnega epitelija se pri žuželkah ne oblikuje, telesna votlina pa postane mešana - mixocoel. Koelomični zametki se združijo s primarno telesno votlino.
Kasneje iz ektoderma nastaneta živčevje in trahealni sistem. Malpigijeve žile nastanejo iz sten zadnjega črevesa.
V procesu razvoja je zarodek žuželke podvržen segmentaciji, ki se najprej manifestira v sprednjem delu, nato pa v zadnjem delu telesa. V predelu glave je akron z očesnimi, labialnimi in

antenske režnje, interkalarni segment in tri čeljustne segmente. Nato nastanejo trije torakalni in deset trebušnih segmentov ter analni reženj.
Pri mnogih žuželkah gre zarodek skozi tri faze, za katere je značilna različna sestava začetkov udov: protopod, polipod in oligopod (slika 339).
Za embrionalni razvoj žuželk je značilen pojav blastokineze. To je sprememba položaja telesa zarodka v jajcu, pri katerem se rezerve rumenjaka najbolj uporabljajo.
Dve vrsti blastokineze za žuželke opisuje A. G. Sharov. Pri žuželkah z nepopolno preobrazbo se zarodek najprej nahaja s hrbtom navzgor in glavo proti sprednjemu koncu jajčeca, nato pa, ko se oblikuje amnijska votlina, se zarodek obrne z ventralno stranjo navzgor in glavo, v tem zaporedju je v zadnjem delu jajca.
Blastokineza poteka različno pri večini žuželk s popolno preobrazbo in pri pravokrilcih, pri katerih se zarodek pogrezne v rumenjak, ne da bi spremenil položaj telesa v jajčecu.
V embrionalnem razvoju žuželk se kažejo prilagoditve na življenje na kopnem: zaščitne membrane (horion, seroza, amnion), zaloga hranil (veliko rumenjaka), amnijska votlina, napolnjena s tekočino.
Pred izvalitvijo oblikovana ličinka žuželke pogoltne tekočino iz amnijske votline, zaradi česar se telesni turgor poveča. Ličinka z glavo prebije horion, ki ima pogosto jajčne zobce ali konico.
Postembrionalni razvoj. V obdobju postembrionalnega razvoja žuželk, po izvalitvi iz jajčeca, pride do rasti mlade živali z zaporednimi molti in prehodom kvalitativno različnih faz razvoja. Med ontogenezo ali individualnim razvojem se žuželke moltajo od 3-4 do 30-krat. V povprečju je število povezav 5-6. Interval med moltami se imenuje stadij, stanje razvoja pa starost. Morfološke spremembe med razvojem od ličinke do odraslega se imenujejo metamorfoza. Pri vseh žuželkah, razen pri nižjih oblikah brez kril, po doseganju odraslega stanja prenehata rast in taljenje odraslih. Zato na primer razlik v velikosti hroščev iste vrste ni mogoče pripisati različnim starostnim skupinam, ampak jih je treba obravnavati le kot manifestacijo individualne variabilnosti.
Obstajajo tri glavne vrste postembrionalnega razvoja žuželk: 1) neposredni razvoj brez metamorfoze - ametabolizem ali protometabolizem; 2) razvoj z nepopolno preobrazbo ali s postopno metamorfozo, - hemimetabolizem; 3) razvoj s popolno preobrazbo, tj. z izrazito metamorfozo - holometabolizem.
Ametabolizem, ali neposreden razvoj, opažamo le pri primarnih brezkrilih žuželkah iz reda ščetinastorepcev (Thysanura), kamor sodi navadna srebrna ribica (Lepisma). Enako vrsto razvoja opazimo pri Entognatha: poskočni rep (Collembola) in dvorepi (Diplura).
Z ametabolizmom se iz jajčeca pojavi ličinka, podobna odrasli osebi. Razlike se nanašajo le na velikost, proporce telesa in stopnjo razvoja spolnih žlez. Za razliko od krilatih žuželk se še naprej talijo tudi v namišljenem stanju.
Hemimetabolizem- nepopolna transformacija ali razvoj s postopno metamorfozo. Značilen je za številne krilate žuželke, na primer za ščurke, kobilice, kobilice, stenice, cikade itd.
S hemimetabolizmom se iz jajčeca pojavi ličinka, podobna odraslim, vendar z osnovnimi krili in nerazvitimi spolnimi žlezami. Takšne odrasle podobne ličinke z zametki kril imenujemo nimfe. To ime je izposojeno iz starogrške mitologije in se nanaša na božanska krilata bitja v obliki deklet. Nimfe žuželk se večkrat linijo in z vsakim taljenjem se njihovi zametki kril povečajo. Starejša nimfa se lini in iz nje nastane krilati imago. Na sliki 340 so prikazane faze razvoja kobilic (jajčeca, nimfe 1.-5. starosti in odrasli) kot primer nepopolne preobrazbe. Ta tipična nepopolna transformacija se imenuje hemimetamorfoza.
Med žuželkami z nepopolno metamorfozo obstajajo primeri razvoja, ko se nimfe opazno razlikujejo od odraslih v prisotnosti posebnih prilagoditev ličink - začasnih organov. Takšen razvoj opazimo pri kačjih pastiricah, majskih muhah, kamenkah. Nimfe teh žuželk živijo v vodi, zato se imenujejo

riž. 340. Razvoj z nepopolno metamorfozo pri kobilici Locusta migratoria (po Kholodkovskem): 1 - protoraks, 2 - mezotoraks s krilnimi popki, 3 - metatoraks s krilnimi popki
najade (vodne nimfe). Imajo začasne organe, kot so trahealne škrge, ki pri kopenskih odraslih izginejo. In ličinke kačjih pastirjev imajo še vedno "masko" - spremenjeno spodnjo ustnico, ki služi za prijemanje plena.
Holometabolizem- popolna preobrazba. Razvojne faze pri holometaboliju so: jajce - ličinka - lutka - imago (slika 341). Ta razvoj je značilen za hrošče, metulje, dvokrilce, himenoptere, tuličarje in čipkarice.

riž. 341. Razvoj s popolno preobrazbo pri sviloprejki Bombyx top (po Leinesu): A - samec, B - samica, C - gosenica, D - zapredek, E - zapredek.
Ličinke žuželk s popolno preobrazbo niso podobne odraslim in se ekološko pogosto razlikujejo. Tako na primer ličinke majskih hroščev živijo v tleh, odrasli pa na drevesih. Ličinke mnogih muh se razvijejo v zemlji, gnijočem substratu, odrasli pa letijo in obiskujejo rože ter se hranijo z nektarjem. Ličinke takšnih žuželk se večkrat linijo in nato spremenijo v lutko. V fazi pupa se pojavi histoliza - uničenje larvalnih organov in histogeneza - nastanek organizacije odrasle žuželke. Iz lutke se pojavi krilata žuželka - imago.
Tako so pri žuželkah opažene naslednje vrste postembrionalnega razvoja: ametabolizem ali protomorfoza (jajce - ličinka (podobno imagu) - imago); hemimetabolija - nepopolna preobrazba (jajce - nimfa - imago): hemimetamorfoza - tipična varianta, hipomorfoza - zmanjšana metamorfoza, hipermorfoza - povečana metamorfoza; holometabolija - popolna preobrazba (jajce - ličinka - lutka - odrasli): holometamorfoza - tipična različica, hipermetamorfoza - z več vrstami ličink.
Vrste ličink žuželk s popolno preobrazbo. Ličinke holometaboličnih žuželk imajo bolj poenostavljeno strukturo v primerjavi z odraslimi. Nimajo sestavljenih oči, rudimentov kril; ustni aparat je griznega tipa, antene in noge so kratke. Glede na razvoj okončin ločimo štiri vrste ličink: protopodi, oligopodi, polipodi in apodi(Slika 342). Ličinke protopoda so značilne za čebele, ose. Imajo le začetke prsnih nog. Te ličinke so neaktivne, razvijajo se v satovju s skrbnostjo delavcev. Ličinke oligopodov so pogostejše od drugih, zanje je značilen normalen razvoj treh parov hojalnih nog. Oligopodi vključujejo ličinke hroščev, čipk. Ličinke polipodov ali gosenice imajo poleg treh parov torakalnih nog še nekaj parov lažnih nog na trebuhu. Ventralna krara predstavlja izbokline trebuha
stene telesa in nosijo trnke in bodice na podplatu. Gosenice so značilne za metulje in žage.Apodične ali breznoge ličinke opazimo v redu Diptera, pa tudi pri nekaterih hroščih (ličinke mrene, vrtalke), metuljih.

riž. 342. Ličinke žuželk s popolno preobrazbo (iz Barnesa): A - protopod, B, C - oligopod, D - polipod, E, F, G - apod
Glede na načine gibanja se ličinke žuželk s popolno preobrazbo delijo na kampodeoid z dolgim, prožnim telesom, tekaškimi nogami in občutkom cerci, erukoid z mesnatim, rahlo ukrivljenim telesom z ali brez okončin, žica- s togim telesom, okroglega premera, s podpornimi cerci - urogomfi in črvasto- brez nog.
Kampodeoidne ličinke so značilne za številne plenilske hrošče - hrošče, hrošče. Premikajo se skozi luknje v tleh. Tipična erukoidna ličinka je ličinka majskega hrošča, hrošča, brona. To so roveče ličinke. Žičniki so značilni za hrošče klikne in temne hrošče, katerih ličinke aktivno delajo prehode v tleh. Številne ličinke, podobne črvom. Premikajo se v tleh, rastlinskih tkivih. Sem spadajo ne le ličinke Diptera, ampak tudi nekateri hrošči, metulji, žage, ki se razvijajo na primer v rastlinskih tkivih.
vrste lutk. Pupae so proste, pokrite in skrite (slika 343). Pri prostih lutkah so zametki kril in okončin jasno vidni in prosto ločeni od telesa, na primer pri hroščih. Pri pokritih lutkah se vsi zametki tesno držijo telesa, na primer pri metuljih. Integumenti prostih mladičev so tanki, mehki, pokritih mladičev pa so močno sklerotizirani.

riž. 343. Vrste lutk pri žuželkah (po Weberju): A - prosti hrošč, B - pokriti metulj, C - skrita muha; 1 - antena, 2 - zametki kril, 3 - noga, 4 - spirale
koža ličinke, ki tvori lažni kokon – puparijo. Znotraj puparija je odprta pupa. Zato je skrita krizalija le različica brezplačne. Puparije so značilne za številne muhe.
Pogosto si ličinka zadnjega stadija pred mladičkom splete kokon. Tako na primer gosenica metuljev sviloprejk izloča svilo iz svilenih žlez, iz katerih prede gost kokon. Znotraj takega kokona je pokrita krizalija. In v nekaterih Hymenoptera - mravljah, kot tudi v Neuroptera, je odprta ali prosta lutka znotraj kokona. Pri ličinkah čipk, kot so zlate oči, nitke za kokon proizvajajo malpigijeve žile in se izločajo iz anusa.
Fiziologija metamorfoze. V procesu metamorfoze potekata dva med seboj povezana procesa: histoliza in histogeneza. Histoliza je razgradnja tkiv organov ličink, histogeneza pa tvorba organov odrasle žuželke. Pri žuželkah z nepopolno metamorfozo se ti procesi odvijajo postopoma v fazi nimfe, pri žuželkah s popolno preobrazbo pa v fazi pupa.
Histoliza nastane zaradi aktivnosti fagocitov in encimov. V tem primeru se najprej uničijo maščobno telo, mišice ličink in nekateri drugi organi, ki se pretvorijo v hranilni substrat, ki ga porabijo tkiva v razvoju.
Histogeneza ali tvorba organov odrasle žuželke se pojavi predvsem zaradi razvoja imaginalnih diskov - zametkov iz nediferenciranih celic. Imaginalni diski se odlagajo v fazi ličinke in celo med embriogenezo ter predstavljajo notranje zametke. Imaginalni diski razvijejo oči, krila, ustni aparat, noge, pa tudi notranje organe: mišice, spolne žleze. Prebavni sistem, malpigijeve žile in sapniki niso uničeni, vendar so v procesu metamorfoze močno diferencirani. Najmanj metamorfizirano srce in živčni sistem. Vendar pa med metamorfozo v živčnem sistemu pogosto opazimo proces oligomerizacije (fuzije) ganglijev.
Proces metamorfoze nadzirajo endokrine žleze (slika 329). nevrosekretorne celice možgani izločajo hormone, ki spodbujajo aktivnost srčna telesa, katerega hormoni prek hemolimfe spodbujajo protorakalni(protorakalne) žleze, ki izločajo talilni hormon - ekdizon. Ecdysone spodbuja proces taljenja: delno raztapljanje in luščenje stare povrhnjice ter nastanek nove.
V procesu metamorfoze aktivnost sosednja telesa proizvajajo mladoletnik hormon. Pri visokih koncentracijah taljenje ličinke povzroči nastanek ličinke
naslednja starost. Ko ličinke rastejo, aktivnost sosednjih teles oslabi in koncentracija juvenilnega hormona pade, protorakalne žleze pa postopoma degenerirajo. To vodi do dejstva, da se ličinke prelijejo v fazo lutke in nato v odraslo osebo.
Umetna presaditev sosednjih teles, na primer v nimfo kobilice zadnje starosti, prispeva k dejstvu, da se ne prelije v odraslo fazo, temveč v večjo ličinko dodatne starosti. V odrasli fazi juvenilni hormon nadzoruje razvoj spolnih žlez, hormon ekdizon pa se ne proizvaja več zaradi zmanjšanja protorakalnih žlez.
Izvor metamorfoze. Obstaja več hipotez o izvoru metamorfoze pri žuželkah. Dolgo časa so potekali spori o tem, katere žuželke so evolucijsko naprednejše - s popolno ali nepopolno preobrazbo. Po eni strani so žuželčje nimfe z nepopolno preobrazbo bolj progresivno razvite kot žuželčje ličinke s popolno preobrazbo, po drugi strani pa imajo slednje napredovalo pupalno fazo.
Trenutno je to protislovje odpravljeno s hipotezo o izvoru metamorfoze G. S. Gilyarova, A. A. Zahvatkina in A. G. Sharova. Po tej hipotezi sta se obe obliki metamorfoze pri žuželkah razvili neodvisno od enostavnejše vrste razvoja - protomorfoze, opažene pri primarnih žuželkah brez kril, na primer pri žuželkah s ščetinami (Thysanura).
Pri protomorfozi je razvoj neposreden, pri čemer opazimo veliko taljenja v fazi ličinke in nato v imaginarnem stanju. Vse faze razvoja teh žuželk potekajo v istem okolju.
Predpostavlja se, da so žuželke v procesu evolucije prešle iz polskritega bivanja v zgornji plasti zemlje v življenje na njeni površini in na rastlinah. Ta prehod v nov življenjski prostor je bil kronan z veliko aromorfozo - razvojem kril in letenjem.
Razvoj odprtih habitatov se je odrazil v evoluciji individualnega razvoja žuželk. Razvoj ontogeneze žuželk je očitno potekal v dveh glavnih smereh.
V enem primeru je potekal proces embrionizacije razvoja, ki je privedel do izleganja žuželk iz jajčec, bogatih z rumenjakom, v kasnejših fazah razvoja. To je vodilo do domišljije ličink z nastankom nimf. Tako so se razvile žuželke z nepopolno preobrazbo. Ta pot evolucije je vodila do progresivnega razvoja ličink, ki vodijo podoben način življenja kot odrasli.
V drugem primeru je, nasprotno, potekal proces deembrionizacije razvoja, to je, da so jajca, revna z rumenjakom, izvaljena v zgodnejših fazah razvoja. To je privedlo do morfoekološkega razhajanja ličink in odraslih žuželk. Ličinke so se poenostavile in prilagodile
živeti v bolj varovanem okolju, opravljati funkcijo prehranjevanja, odrasli pa so začeli opravljati predvsem funkcijo razmnoževanja in naselitve. Poleg deembrionizacije razvoja ličink žuželk s popolno preobrazbo so razvile številne začasne prilagoditve na različne pogoje obstoja. Tako je prišlo do amfigeneze (divergenca) v evoluciji ličink in odraslih žuželk s popolno metamorfozo. Amfigeneza ličink in odraslih se je izkazala za zelo globoko v smislu morfoloških prilagoditev, kar je povzročilo resna protislovja v ontogeniji. Uspešno so jih rešili s pojavom pupalne faze, med katero pride do korenitega prestrukturiranja larvalne organizacije v imaginalno. To je omogočilo žuželkam s popolno preobrazbo, da obvladajo širšo paleto ekoloških niš in dosežejo razcvet brez primere med živalmi na Zemlji.
Gojenje žuželk. Za večino žuželk je značilno biseksualno spolno razmnoževanje. Mnoge vrste kažejo spolni dimorfizem. Samci hroščev jelenov imajo na primer mandibule spremenjene v rogove, samci hroščev nosorogov pa imajo rog na glavi in grbe na pronotumu. To je posledica paritvenega vedenja teh vrst, ki ga spremlja boj samcev za samico. Razmerje med spoloma pri različnih vrstah je izjemno raznoliko. Samci Diptera iz družine Dolichopodidae prinesejo samici "darilo" - ujeto muho in zaplešejo z ogledali na nogah. Samice bogomolke so plenilske in med parjenjem pojedo samca.
Večina žuželk odlaga jajca, vendar pogosto opazimo živo rojstvo. V tem primeru se jajčeca razvijejo v ženskem genitalnem traktu in rodi ličinke. Na primer, sarkofagi (Sarcophagidae) odlagajo žive ličinke na meso, katerih razvoj poteka zelo hitro. Ni čudno, da so v starih časih verjeli, da črvi v mesu nastanejo spontano. Lahko ne opazite, kako je mehurček obiskal odprto ležeče meso, in nenadoma odkrijete bele ličinke, ki so se nenadoma pojavile.
Med živorodne vrste spadajo tudi muha ovčja krvosesa, nekateri hrošči, ki živijo v jamah.
Poleg dvospolnega spolnega razmnoževanja številne žuželke izkazujejo partenogenezo – razvoj brez oploditve. Obstaja veliko vrst iz različnih redov žuželk, za katere je značilna partenogeneza. Partenogeneza je lahko obvezna - obvezna, takrat so vsi posamezniki vrste samo samice. Tako v alpskih razmerah, na severu in v drugih neugodnih razmerah najdemo partenogenetske hrošče, pravokrilce, ušesce in čipke. Partenogeneza se pojavlja tudi pri dvospolnih vrstah, ko je del jajčec odloženih oplojenih, del pa neoplojenih. Čebelji troti se na primer razvijejo iz neoplojenih jajčec.
Podobno partenogenezo najdemo tudi pri drugih Hymenoptera (mravlje, žagice), termitih, nekaterih stenicah in hroščih. In pri listnih uših, na primer, pride do spremembe generacij v življenjskem ciklu: biseksualne in partenogenetske. V nekaterih primerih je partenogeneza lahko neobvezna (začasna) in se manifestira le v neugodnih pogojih. Partenogeneza pri žuželkah prispeva k ohranjanju visokih populacij.
Različica partenogeneze je pedogeneza - razmnoževanje brez oploditve v larvalni fazi razvoja. To je poseben način razvoja žuželk, ko je zorenje spolnih žlez pred drugimi organi. Na primer, nekatere vrste žolčnih mušic se razmnožujejo v fazi ličink. Starejše ličinke skotijo mlajše ličinke. Pedogenezo so opazili pri eni od vrst hroščev, katere ličinke delno odlagajo jajčeca in delno rodijo ličinke. Pedogeneza poleg dvospolnega razmnoževanja povečuje številčnost vrste.
Življenjski cikel žuželk. Za razliko od ontogeneze oziroma individualnega razvoja žuželk je življenjski cikel razvoj vrste, ki praviloma vključuje več tipov ontogenij. Ontogeneza je omejena na življenje enega posameznika od jajčeca do začetka pubertete in nato naravne smrti. Življenjski cikel je ponavljajoči se del nenehnega razvoja vrste. Tako je v najbolj tipičnem primeru pri žuželkah življenjski cikel sestavljen iz dveh konjugiranih in morfofiziološko različnih ontogenij samcev in samic, ki se spolno razmnožujejo in razmnožujejo sebi podobne. In pri partenogenetskih vrstah je za življenjski cikel značilna samo ena vrsta ženske ontogeneze.
Življenjski cikli žuželk so raznoliki glede na vrste razmnoževanja, sestavo generacij in njihovo menjavanje. Razlikujemo lahko naslednje vrste življenjskih ciklov žuželk.
1. Življenjski cikli brez menjave generacij z biseksualnim spolnim razmnoževanjem. To je najpogostejši tip življenjskega cikla, ki je značilen za dimorfne vrste, ki jih sestavljajo samo samci in samice, ki se spolno razmnožujejo. To so cikli večine hroščev, metuljev, stenic.
2. Življenjski cikli brez menjave generacij s partenogenetskim razmnoževanjem. Takšne vrste so monomorfne, sestavljene so samo iz partenogenetskih samic, ki odlagajo jajčeca brez oploditve.
Partenogenetske vrste so še posebej pogoste med listnimi ušmi, luskarji in drugimi enakokrilci. V visokogorju so pogoste partenogenetske vrste hroščev, hroščev, kobilic in kokic.
3. Najredkejša vrsta življenjskega cikla pri žuželkah je cikel brez menjave generacij s spolnim razmnoževanjem hermafroditnih vrst.
Znana je ameriška vrsta mušnice, ki jo sestavljajo samo hermafroditski posamezniki. V zgodnjih fazah razvoja odrasli delujejo kot samci, v poznejših fazah pa kot samice. Zato vsi posamezniki odlagajo jajca, kar poveča številčnost vrste.
4. Življenjski cikli brez menjave generacije s spolnim razmnoževanjem in fakultativno partenogenezo pri polimorfnih vrstah, na primer pri družbenih žuželkah. Vrsto sestavljajo spolni posamezniki - samci in samice ter plodni - delovni posamezniki, ki ne sodelujejo pri razmnoževanju. Te vrste vključujejo čebele, mravlje in termite. Takšne življenjske cikle otežuje dejstvo, da samice poleg oplojenih jajčec odlagajo tudi partenogenetske, iz katerih se na primer čebele razvijejo haploidni samci - troti, iz oplojenih pa samice in delavke. Nekateri jahači, trips, kokcidi se razvijajo podobno.
Pri drugih vrstah se fakultativna partenogeneza kaže na drugačen način: iz neoplojenih jajčec se razvijejo ne samci, ampak samice. Toda v tem primeru se diploidni nabor kromosomov pri ženskah obnovi s fuzijo haploidnih jeder. Takšen razvoj poznamo pri nekaterih paličnjakih, kobilicah, žagicah, kokcidih.
5. Življenjski cikli z menjavo spolne generacije in partenogenetske (heterogonije). Pri mnogih listnih uših in filokseri je poleg spolne generacije krilatih samcev in samic več izmeničnih generacij partenogenetskih samic, krilatih ali brez kril.
6. Življenjski cikli z menjavo spolne generacije in več generacij s pedogenezo. Na primer, pri nekaterih žolčnikih po spolnem razmnoževanju, pri katerem sodelujejo samci in samice, pride do partenogenetske reprodukcije ličink (pedogeneza). Po več generacijah plemenskih ličink, ki po rojstvu lastne vrste odmrejo, se zadnja generacija ličink zabubi in povzroči krilate samice in samce.
7. Življenjski cikli z menjavo spolne generacije (moški in samice) z nespolnimi. Po spolnem razmnoževanju samice odložijo oplojena jajčeca, ki so podvržena poliembrioniji. To je nespolno razmnoževanje v embrionalni fazi. Jajčece se zdrobi in zarodek v fazi morule se začne razmnoževati z brstenjem. Iz enega jajčeca lahko nastane več deset zarodkov. Takšna
Tako lahko razvrstitev življenjskih ciklov predstavimo na naslednji način.
JAZ. Brez menjave generacij:
- 1) z biseksualnim spolnim razmnoževanjem (majski hrošč);
- 2) s partenogenetskim razmnoževanjem (alpski hrošči, kobilice);
- 3) s spolnim razmnoževanjem hermafroditnih posameznikov (ameriška muha);
- 4) s spolnim razmnoževanjem in delno partenogenezo pri polimorfnih vrstah (čebelah).
II. Z menjavo generacij:
- 1) heterogonija: menjava spolne generacije in več partenogenetskih (listne uši, filoksera);
- 2) heterogonija: menjava spolne generacije in več pedogenetskih generacij (nekatere žolčniki);
- 3) metageneza: menjava spolne generacije s poliembrionijo (jezdeci).
Sezonski cikli žuželk. Če življenjski cikel razumemo kot ciklično ponavljajoč se del morfogeneze vrste iz ene razvojne faze v isto fazo, potem sezonski razvojni cikel razumemo kot značilnost razvoja vrste v letnih časih enega leta ( iz zime v zimo).
Tako na primer življenjski cikel majskega hrošča traja 4-5 let (od jajčeca do zrelih odraslih osebkov), za sezonski cikel te vrste pa je značilno, da se prezimljene ličinke spomladi zabubijo in mladi hrošči razmnožujejo. Poleti, jeseni in pozimi najdemo njihove ličinke različnih starosti. Število generacij, ki se razvijejo med letom, se imenuje voltnost.
Obstajajo vrste, ki dajejo več generacij na leto. To so multivoltaične vrste. Na primer, hišna muha lahko daje 2-3 generacije na sezono in prezimi v odrasli fazi. Večina žuželk je monovoltinskih, ki proizvedejo eno generacijo na leto.
Za sezonske cikle žuželk v naravi so značilna koledarska obdobja pojavljanja različnih faz razvoja. Pomembne značilnosti sezonskih ciklov vrst so čas njihovega aktivnega življenja in diapavza (začasna zamuda v razvoju) pozimi ali poleti. Regulacijo življenjskih ciklov vrst v skladu z lokalnimi sezonskimi pojavi zagotavljajo okoljski dejavniki in nevrohumoralni sistem telesa.
Embrionalni ali embrionalni razvoj čebele delavke, maternice, trota vključuje vse spremembe, ki se zgodijo pod jajčno lupino, zaradi česar iz enoceličnega jajčeca nastane večcelično bitje, ličinka (slika 32. ). Jajce, ki ga je pravkar položila maternica, je praviloma z enim koncem prilepljeno na dno celice in stoji navpično na njem. Dolžina jajčeca je 1,6-1,8 mm, širina 0,31-0,33 mm. Je rahlo ukrivljen, podolgovate cilindrične oblike, konec nasproti mesta pritrditve je rahlo razširjen. Vklopljeno
- 3. dan jajčece zavzame nagnjen položaj, 3. dan pa leži na dnu celice. Na prostem razširjenem koncu jajčeca je drobna luknjica, skozi katero vstopi semenčica iz maternične posode med prehodom jajčeca skozi jajcevod. Ta luknja se imenuje mikropil. V drugih primerih je mikropil neprepusten za bakterije in viruse. Jajce ima sprednji in zadnji konec. Glava ličinke se razvije v prostem sprednjem koncu jajčeca.
Embrionalni razvoj se začne z drobljenjem jedra. Hčerinska jedra, ki nastanejo kot posledica delitve, se najprej naključno nahajajo znotraj rumenjaka, nato pa se z majhnimi deli citoplazme premaknejo na obrobje jajčeca in tvorijo neprekinjeno plast celic - blastodermo, ki obloži celotno jajčno lupino od notranjost. Na konveksni strani jajčeca začnejo celice blastoderma rasti in se množiti hitreje kot celice na konkavni strani. Posledično se oblikuje embrionalni trak v obliki širokega pramena. To je začetek nastajanja zarodka. Zarodni vod raste zaradi delitve in diferenciacije celic, nastajanja tkiva. Kot rezultat teh procesov pride do razvoja ličinke. V drugi polovici 2. dneva se začne proces segmentacije zarodka. Na sprednjem koncu zarodnega traku se oblikuje predoralni reženj (akron), na katerem se oblikuje tuberkel - zametek zgornje ustnice. Pod tem tuberkulom se pojavi vdolbina - bodoča usta. Nato - zametki anten. Segmenti prsnega koša in trebuha se odcepijo od segmenta glave nazaj. Na ventralni (ventralni) strani teh segmentov se pojavijo izrastki - zametki seznanjenih okončin. Najprej se pojavi na prsih, kasneje pa na trebuhu. Hkrati se v predelu glave pojavi izrazita segmentacija: nastanejo zametki petih segmentov glave. Poleg tega se pojavijo dihalne luknje - stigme, pa tudi začetki predilnih žlez in Malpigijevih žil. Ko se oblikujejo segmenti telesa in okončin, pride do ločevanja posameznih delov.
Kasneje se zarodna črta poveča. Razmnoževanje blastodermnih celic vodi do invaginacije srednjega dela zarodnega traku (proces gastrulacije), ki se spusti globlje v jajčece in se loči od blastoderma. V procesu gastrulacije se na površini zarodnega traku najprej oblikuje vzdolžni žleb, nato pa začneta zunanja robova žleba rasti drug proti drugemu. Kot rezultat, bodoča druga zarodna plast invaginira znotraj zarodnega traku. Ko se robovi žleba zaprejo, nastane zunanja plast (ektoderm). Pod njo leži notranja plast (mezoderm). Na koncih mezodermalne plasti se pojavijo invaginacije, sestavljene iz skupin celic, ki tvorijo tretjo zarodno plast (endodermo). Začnejo se pojavljati zametki posameznih organov. Njihov razvoj in diferenciacija se nadaljuje, dokler se iz celice ne izleže ličinka.
Iz ektoderma nastane telesna stena, sprednje in zadnje črevo, sapnik in živčni sistem ter številne žleze; iz endoderma - zametki žela, kril in okončin, končni deli genitalnega trakta, živčni sistem in čutilni organi; iz mezoderma - mišice, maščobno telo, spolne žleze.
Popolnoma oblikovan zarodek, ki se je v bistvu spremenil v ličinko, zapolni celotno jajce. Začne se intenzivno gibati, vleči zrak v sapnik, požirati amnijsko tekočino, s čimer poveča prostornino svojega telesa. Ličinka se sprosti iz embrionalnih ovojnic, zlomi horion (sekundarna lupina jajčeca, ki nastane s transformacijo folikularnih epitelijskih celic) in se izleže, to je, da pride ven. Običajno embrionalni razvoj medonosnih čebel traja približno 3 dni. Spremembe temperature lahko podaljšajo ali skrajšajo obdobje razvoja zarodka.
Po sprostitvi ličinke iz jajčeca se začne postembrionalni razvoj, ki ga spremlja metamorfoza (preobrazba).
Metamorfoza je globoka transformacija v strukturi organizma, zaradi katere se ličinka spremeni v odraslo žuželko. Glede na naravo postembrionalnega razvoja pri žuželkah ločimo dve vrsti metamorfoze:
nepopolna (hemimetabolizem), ko je za razvoj žuželke značilen prehod le treh stopenj - jajčeca, ličinke in odrasla faza (odrasla);
popoln (holometabolizem), ko se prehod ličinke v odraslo obliko izvede na vmesni stopnji - stopnji lutke.
Z nepopolno preobrazbo je način življenja zgodnjih faz in odraslih podoben. Ličinka s to vrsto razvoja je podobna odrasli žuželki in ima tako kot ona sestavljene oči, ustne organe in nato zametke kril, ki so jasno vidni od zunaj. Rudimenti kril se z vsakim naslednjim moltom povečujejo in v zadnji predimaginalni dobi lahko že pokrivajo več segmentov trebuha. Pri žuželki zadnje predimaginalne dobe so imaginalne strukture popolnoma razvite, odrasel pa se pojavi kot posledica zadnjega molta v celoti oblikovan. Nepopolna preobrazba je značilna za pravokrilce, stenice, homoptere itd.
S popolno metamorfozo je za celoten razvojni cikel značilen prehod stopenj jajčeca, ličinke, predlečke, lutke in odraslega (tabela 3).
- Trajanje stopenj razvoja posameznikov čebelje družine, dni
Prehod iz ličinke v odraslo osebo poteka v fazi lutke. To je nehranilna faza, med katero pripadajoča tkiva
V zgodnejših fazah se oblikujejo imaginarne strukture. Pri večini žuželk je ta stadij negibljiv. Pupa se ne odziva na zunanje dražljaje in je oblečena v gosto prevleko. Na koncu te stopnje lupina poči in pojavi se odrasla žuželka.
Pri žuželkah regulacijo metamorfoze izvajajo hormoni, ki jih proizvajajo endokrine žleze (endokrine žleze). Trenutno so znani trije hormoni metamorfoze, ki nadzorujejo postembrionalni razvoj: juvenilni hormon, hormon moltinga (ličinke) in aktivacijski hormon. Juvenilni hormon se proizvaja in shranjuje v sosednjih telescih, ki sta dve majhni celični strukturi, ki ležita za možgani na straneh požiralnika.
Če mladi ličinki odstranimo sosednja telesca, bo mladiček sledil naslednjemu moltu, kljub dejstvu, da bi moral v normalnem stanju narediti še nekaj moltov. Ta hormon določa rast ličink, nadzoruje razvoj številnih organov in procesov. Na primer, maščobno telo ne raste in ne deluje brez tega hormona.
Hormon taljenja ali ekdizon nastaja v protorakalnih žlezah, ki se nahajajo v protoraksu ličinke nad prvim živčnim ganglijem blizu prvega dihalca. Sintetizira se v telesu žuželk iz. holesterol. Hormon sproži proces taljenja in tako posredno uravnava rast in nastanek strukture. Vnos tega hormona žuželkam povzroči nastanek oteklin na velikanskih kromosomih. Analiza teh oteklin je pokazala, da v njih poteka intenziven proces tvorbe RNK. Domneva se, da je prvi rezultat delovanja ekdizona aktivacija genov, nato stimulacija biosinteze RNA in tvorba ustreznih encimov, ki zagotavljajo proces metamorfoze.
Aktivacijski hormon nastaja v posebnih nevrosekretornih celicah, ki se nahajajo v sprednjem dorzalnem delu supraezofagealnega ganglija. Bolje so te žleze razvite pri čebelah delavkah, manj pri maticah in še manj pri trutih. Aktivacijski hormon vpliva na ponovni začetek aktivnosti žuželk po vsakem taljenju in spodbuja tvorbo dveh drugih hormonov - ecdysone in juvenile.
Medonosna čebela je žuželka s popolno preobrazbo (slika 33).
Stadij ličinke. Zanj je značilna intenzivna prehrana in rast. Stadij ličinke delavske čebele traja 6 dni, kraljica - 5, dron - 7. V prvih dneh ličinke medonosnih čebel prejmejo toliko hrane, da plavajo na njeni površini. Osnova krme je skrivnost hipofarina-
| 1 g | 3 | 4 5 | 6 7 8 amperov; | 10 | 11 12 | 13 14 15 16 17 18 19 20 | 21 |
| w |
Dan
riž. 33. Faze razvoja čebele
žleze čebele dojilje - prosojna bela tekočina. Mleko, ki ga izločajo žleze čebele dojilje, je zelo hranljivo. Vsebuje ogljikove hidrate, beljakovine, maščobe, minerale, vitamine skupine B. Preden da ličinki hrano, čebela delavka, ki spusti glavo v celico, večkrat odpre in zapre zgornje čeljusti. Nato se s čeljustmi dotakne dna celice in sprosti tekočo hrano. Druge čebele dojilje na enak način dodajajo naslednje porcije mleka v celico z ličinko. Za celotno fazo ličinke je približno 10 tisoč obiskov celice z eno ličinko.
Hrana mladih ličink in starejših ličink ni enaka. V drugem obdobju življenja začne ličinka čebele delavke prejemati kašo - mešanico, ki jo čebele dojilje pripravijo iz medu in cvetnega prahu.
V fazi ličinke se linearne dimenzije ličink delavk povečajo za več kot 10-krat. Tako je dolžina novo izležene ličinke 1,6 mm, 1 dan stare ličinke 2,6 mm, 2 dni stare ličinke 6 mm, do konca ličinke pa 17 mm (skoraj v celoti zavzame dno celice). Dolžina ličinke maternice do konca faze doseže 26,5 mm.
Intenzivna prehrana zagotavlja ličinkam znatno povečanje telesne teže. Izležena ličinka tehta približno 0,1 mg. Za prvo
- dni se masa ličinke delavske čebele poveča za 45-krat, masa ličinke trota pa za 85-krat. Do konca faze se teža ličink delovnih osebkov poveča za 1565-krat, mater pa za 2926-krat.
Po videzu se ličinka močno razlikuje od odrasle čebele (slika 34). Ima črvasto obliko, mehke pokrove belkaste barve. Njegovo telo je sestavljeno iz glave, členkastega trupa in analnega režnja.
Glava ličinke je majhna, tupo stožčasta, osnovo stožca tvori kapsula glave, ki je razdeljena z vzdolžnim šivom.
riž. 34. Zunanja (G) in notranja (1G) zgradba čebelje ličinke "\u003e
riž. 34. Zunanja (D) in notranja (1D) zgradba čebelje ličinke:
A - splošni pogled; B - pogled s strani glave; C - položaj odrasle ličinke na dnu celice; G - stranski pogled; y - zametki anten; vg - zgornja ustnica; vh - zgornja čeljust; opzh - odprtina predilne žleze; ng - spodnja ustnica; p - usta; ndg - supraezofagealni ganglij; PC - predželuce; a - aorta; sk - srednje črevo; zht - maščobno telo; zpzh - zametek spolne žleze; c - srce; zk - zadnje črevo; ms - malpigijeva posoda; bn - trebušna živčna veriga; pzh - predilni likalnik; pdg - subezofagalni ganglij
na dva enaka konveksna dela, tako imenovana lica. Vrh stožca na sprednjem delu glave tvorijo zgornja ustnica in drugi ustniki. Del glave, vključno s spodnjo ustnico, zgornjo čeljustjo in delom glavične kapsule, na katerega so pritrjeni, je ločen od glavične kapsule z globokim utorom, ki poteka na vsaki strani med dnom zgornje čeljusti in maksilo ter se razteza do zadnjih robov glave.
Mandibule segajo iz kapsule glave v obliki stožčastih organov. Antenalni rudimenti se nahajajo na kapsuli glave nad dnom mandibul v obliki okroglih tuberkulozov.
Ličinke na prsnih segmentih nimajo zunanjih rudimentov niti kril niti nog. Ni sestavljenih oči. Vendar pa so pod larvalno kožico nakazani njihovi zametki, ki se imenujejo imaginarni popki. Namišljeni popki nog so vretenasti in razrezani s prečnimi žlebovi.Zametki kril so sestavljeni iz dveh ravnih, navznoter upognjenih procesov hipodermisa.Nahajajo se na straneh drugega in tretjega prsnega segmenta.V regiji osmega in devetega trebušnega segmenta na ventralni strani se pojavijo zametki spolnih organov.
Treba je opozoriti, da se razvoj kril, anten, oči in nog ne pojavi s prestrukturiranjem zmanjšanih struktur ličink; razvijejo se iz izoliranih območij embrionalnih tkiv, ki so v neaktivnem stanju, imenovanih zametki.
Ličinke se od odrasle čebele razlikujejo po zgradbi notranjih organov (glej sliko 34). Osrednje mesto v telesu ličinke zavzema črevesni kanal, ki je sestavljen iz treh delov - sprednjega, srednjega in zadnjega črevesa. Sprednje črevo je videti kot kratka tanka cev, ki je na zadnjem koncu rahlo razširjena. Sprednje črevo vsebuje majhno ustno votlino, kratko žrelo in požiralnik. Medena golša, značilna za odrasle čebele, je odsotna. V steni predželudca so mišice, ki zagotavljajo zaužitje tekoče hrane. Na stičišču požiralnika in srednjega črevesa je majhna obročasta guba, ki igra vlogo ventila. Zapira lumen požiralnika in preprečuje, da bi vsebina srednjega črevesa zatekla nazaj v požiralnik. Do konca faze ličinke se namesto zaklopke oblikuje debel septum, ki pokriva srednje črevo. 1
Srednje črevo je največji del črevesja in zavzema večino telesa ličinke. Posebnost tega oddelka je odsotnost njegovih povezav z zadnjim črevesjem, zaradi česar neprebavljeni deli hrane ostanejo v njem skozi celotno fazo ličinke. Povezava teh dveh delov se pojavi šele pred predenjem kokona, ko je ličinka že zaprta. V tem obdobju se membrana, ki ločuje srednje črevo od zadnjega črevesa, prebije in blato preide v zadnje črevo, nato pa se odstrani iz telesa na dno celice.
V srednjem črevesu imajo ličinke malpigijeve žile. Čebela jih ima štiri. So dolge, rahlo zavite cevi, ki potekajo vzdolž srednjega črevesa. Med življenjem ličinke so malpigijeve žile zaprte. Šele do 6. dneva ličinke, ko močno nabreknejo zaradi tam nakopičenih produktov razpadanja, se zadnji konci in vsebina prebijejo, preidejo v zadnje črevo in se nato odstranijo. Tako ličinka med rastjo in hranjenjem ne iztreblja.
Zadnje črevo je majhno in je ozka cev, ukrivljena pod ostrim kotom in se konča v anus.
Srce ličinke je sestavljeno iz 12 komor enake strukture (pri odrasli čebeli jih je le pet) in je tankostenska cev, ki se nahaja neposredno pod povrhnjico trebuha in prsnega koša. V drugem segmentu torakalne regije poteka aorta. Ima videz cevi, upognjene navzdol, ki nato preide v glavo in se konča na sprednji površini možganov, kjer se hemolimfa izliva v telesno votlino. Na površini aorte je gosta mreža sapnikov.
Med vsemi srčnimi komorami so odprtine, skozi katere hemolimfa vstopa v srce. Zaradi dejstva, da tanki sprednji konci vsake komore štrlijo v naslednjo komoro, povratni tok tekočine ni dovoljen.
Dihalni sistem ličinke je precej preprost in ga predstavljata dva velika vzdolžna trahealna debla, ki potekata vzdolž stranic telesa, z majhnimi vejami od njih, ki se širijo po telesu. Na straneh segmentov je 10 parov spiral, ki so vdolbine, obdane z ozkimi hitinastimi obroči.
Živčni sistem je sestavljen iz dveh preprosto razporejenih glavnih ganglijev (velikega supraezofagealnega in majhnega subglotalnega) in ventralne živčne vrvice, ki se razteza vzdolž spodnje strani prsnega koša in trebuha. Trebušna veriga je sestavljena iz 11 ganglijev - 3 torakalnih in 8 trebušnih. Zadnji trebušni ganglij je na sredini enajstega segmenta; nastala je z zlitjem ganglijev zadnjih treh segmentov. Gangliji so med seboj povezani s parnimi povezavami. Subezofagealni ganglij je prav tako povezan z ventralno živčno vrvico preko dveh veziv.
Genitalni organi so prisotni v ličinki čebele delavke v povojih in so predstavljeni z dvema grebenoma, ki se nahajata v osmem segmentu. Njihova dolžina je v povprečju 0,27 mm, širina pa 0,14 mm. V celotnem stadiju ličinke poteka razvoj primordijev jajčnikov. Pri 2-dnevni ličinki so nepomembne velikosti, pri 6-dnevni ličinki so že podolgovate in se nahajajo proti repnemu koncu skozi deveti segment. Do konca ličinke doseže število jajčec v delavski čebeli 130-150. V kasnejših fazah, ko se ličinka spremeni v lutko, tkiva ličinke razpadejo in število jajčec pri čebeli delavki se zmanjša z 20 na 3, medtem ko se v maternici jajčniki še naprej razvijajo v fazi lutke.
Maščobno telo je v ličinki dobro razvito, kjer se kopičijo hranila. Pri starejših ličinkah maščobno telo doseže 60 % telesne teže in zapolnjuje skoraj celotno votlino med srednjim črevesjem in kutikulo ličinke. Maščobno telo je koncentracija maščob, beljakovin in ogljikovih hidratov, zaradi katerih se v prihodnosti razvije pupa. Maščobno telo poleg samih maščobnih celic sestavljajo tudi izločevalne celice in enociti, ki lovijo soli sečne kisline.
Predilna žleza je prisotna le pri ličinkah. Izloček te žleze služi kot material za predenje kokona pred prehodom ličinke v stadij lutke. Predilna žleza je videti kot dve dolgi cevi, ki sta z ventralne strani nameščeni pod srednjim črevesjem. V sprednjem čagu sta obe cevi povezani v neparni izločevalni kanal, ki se odpira pod ustno odprtino na konici spodnje ustnice.
V celotnem stadiju ličinka razvije zametke žela, od katerih se en par nahaja na enajstem segmentu in dva para na dvanajstem. Med njimi po segmentih je zametek genitalne odprtine. Rudiment velike strupene žleze se pojavi šele proti koncu stadija ličinke, na začetku stadija lutke pa se iz poglobitve pokrova na devetem segmentu oblikuje majhna strupena žleza.
5-6 dan po sprostitvi ličink iz jajčeca začnejo čebele celico zapirati z voščenim pokrovčkom. Poleg voska sestava materiala, ki sestavlja kapico, vključuje cvetni prah, vodo in papirno maso. Pokrov se izkaže za porozen, zaradi česar zrak, potreben za dihanje ličinke in nato lutke, prodre v celico. Od tega trenutka ličinka konča svojo rast in se preneha hraniti.
Takoj ko se celica zapre, se ličinka v njej zravna, črevesne stene se skrčijo in neprebavljeni ostanki hrane, ki so najprej prebili tanko steno srednjega črevesa, preidejo v debelo in od tam navzven. Ličinka odloži iztrebek v enega od vogalov celice. Po tem ličinka prede kokon, t.j. plete notranjo površino celice. Pri predenju kokona ličinka uporablja izloček predilne žleze, malpigijeve žile in lepljive izločke iz sten svojega telesa. Postopek predenja je podrobno preučil in opisal Velich (1930).
Med vrtenjem kokona ličinka izvaja hitre ritmične drhteče gibe; začenši od glave, se val premika skozi segmente. V 1 minuti ličinka naredi do 280 gibov. Hkrati s tresenjem ličinka izvaja zibajoče gibe glave s sprednjim delom telesa. So črtkane, eliptične, nepravilne krivulje od zgoraj navzdol ali od desne proti levi. Pri tem iz odprtine z zaklopko na spodnji ustnici priteče prozorna snov, ki jo ličinka izloča v obliki ene debele ali več tankih niti. V 2 dneh ličinka, ki se obrača, krepi stene kokona. Pri predenju kokona so se ličinke čebel delavk, trotov in matic obnašale podobno, razlika je bila le v obliki kokona. Odvisno je od velikosti celice, v kateri se nahaja ličinka. Na primer, pri čebeli delavki kokon ustreza obliki šesterokotne prizme, katere dno je tristrana piramida, pokrov pa natančen odtis zaprte čebelje celice. Kokon po izhodu mlade čebele se ne odstrani. Ker je v eni celici prikazanih več deset generacij, se premer celice sčasoma zoži. Torej, če ima običajna celica sveže zgrajenega satja prostornino 0,282 cm3 in debelino dna 0,22 mm, potem se po 20 generacijah čebeljih osebkov kubatura celice zmanjša na 0,248 cm3, dno pa se odebeli na 1,44 mm. .
Ličinka čebele delavke in matice prede kokon 2 dni, ličinka trota pa 3 dni. Nato se lini še petič. Pred to litvijo sledijo številne spremembe v organizmu ličinke. Rudimenti vseh dodatkov, ki se nahajajo pod kožico, se začnejo obračati navzven ali premikati navzven in se spremeniti v zunanje dele telesa. Ti organi postopoma prevzamejo obliko, značilno za pupa. Glava, ustni deli in noge se obrnejo iz sprednjega v zadnji trebušni del. Po vrtenju kokona se ličinka zravna in zamrzne. Začne se pripravljalni proces na fazo lutke, ki zahteva precej časa. To obdobje imenujemo predpupalna faza.
Predpupalna stopnja. Trajanje faze pred lutko za delavsko čebelo je 3 dni, za kraljico - 2, za drone - 4 dni. Za to obdobje so značilne pomembne spremembe v strukturi ličinke. Ličinka se pod vplivom posebnega talilnega hormona ekdizona popolnoma loči od lupine. Izjema so trahealne povezave med ličinkami in pupalnimi spiralami. Glava je že 2/3 velikosti glave odraslega. Oči se povečajo, njihova površina postane nagubana in nagubana. Prsni koš je od glave ločen z jasno vidno zožitvijo. Krila začnejo rasti v širino, na tej stopnji so nagubana in tesno pritisnjena na prsni koš. Ustni deli se podaljšajo in združijo v strnjeno skupino.
Segmenti prsnega koša in trebuha so razdeljeni vzdolž vodoravne črte na dve polovici - hrbtno in trebušno, ki postopoma pridobivajo strukturo, značilno za odraslega.
Poleg tega so notranji organi podvrženi razpadu. Ta proces se imenuje histoliza. Spremlja ga prodiranje in vnos v tkiva krvnih celic - fagocitov. Viri energije, potrebni za kemične reakcije, na katerih temeljijo ti procesi, so zaloge maščob, glikogena in drugih ogljikovih hidratov v maščobnem telesu ličinke, sladkorja hemolimfe in mišic. Med histolizo začnejo hemociti delovati kot požiralne celice, tj. fagociti, encimi pa pretvorijo maščobno telo, večino mišic ličinke, v hranilni substrat, ki ga hemolimfa dostavlja rastočim tkivom. V poznejših fazah histolize fagocite prevzamejo nova tkiva v razvoju. Mišični sistem je podvržen največjim spremembam, zaradi česar je predpupa imobilizirana.
V sprednjem črevesju se pojavi medena golša z zaklopko, srednje črevo postane zankasto in pridobi nagubano strukturo. V zadnjem črevesju se pojavita dva dela - tanko črevo in rektum. Poveča se število tubulov malpigijevih žil. Opazimo delno zlitje ganglijev nevralne verige, zaradi česar ima lutka sedem ganglijev trebušne verige namesto 11 ganglijev ličinke. Mastno telo se zmanjša.
Stadij pred lutko se konča z odpadanjem stare kutikule in izpod nje se pojavi lutka.
Stadij lutke. Celotna površina telesa novonastale lutke je prekrita z gubami, vendar ni nagubana. Po svoji zgradbi je lutka podobna odrasli čebeli, čeprav je za razliko od slednje nepremična, njeno telo je brez pigmentacije (belo). Vendar se notranji življenjski procesi ne upočasnijo, ampak potekajo z veliko učinkovitostjo. Pupa takoj po zaključku zadnjega molta je prekrita s kožico. Za fazo lutke so značilne globoke preobrazbe. Vir plastičnih snovi za tvorbo novih organov in tkiv ali za histogenezo so razpadni produkti, ki jih prenašajo hemociti. Predilne žleze, prebavni kanal, maščobno telo in mišice popolnoma razpadejo.
Namesto razpadlih organov in tkiv ličinke se oblikujejo novi organi odrasle žuželke.
Glava je veliko daljša in razširjena skoraj do velikosti glave odraslega. Med razširjeno glavo in zoženim protoraksom je most. Ni pa ga med torakalnim in trebušnim segmentom. Pupa še nima kril, vendar so antene, proboscis in noge že jasno označene.
Nekateri elementi maščobnega telesa so v tem obdobju razpršeni v glavi in prsih, nekateri pa so koncentrirani v trebuhu. S starostjo so elementi maščobnega telesa bolj enakomerno razporejeni v glavi, prsih in trebuhu. Struktura želodca se spreminja.
Srce se malo spremeni: število komor se zmanjša z 12 na pet. V celotni fazi lutke srce ne preneha delovati, saj proces metamorfoze zahteva gibanje hemolimfe. Masa supraezofagealnega ganglija v glavi se poveča. To se zgodi zaradi nediferenciranih živčnih celic. Poleg tega se združijo živčni gangliji v prsnem košu, pa tudi zadnji štirje gangliji na koncu trebuha.
Začetki spolnih žlez niso uničeni. Njihova rast v začetni fazi je povezana z razpadom celic maščobnega telesa in sproščanjem plastičnega materiala v obliki albumidov. Ko se razvijejo, pridejo v stik z deli gonadnih kanalov, ki izhajajo iz ektoderma.
Eden od pokazateljev notranjega življenja krizale je njeno dihanje.
Dihalni sistem je med histolizo in histogenezo podvržen številnim spremembam. Torej se prva in tretja torakalna spirala povečata, druga pa se, nasprotno, zmanjša. Uničeno, sapnik. Nove sapnice, pa tudi zračne vrečke, nastanejo iz namišljenih rudimentov, ki se nahajajo na sapničnih deblih. 1
Sestavljene oči rastejo iz popkov na straneh glave. Kot že omenjeno, je lutka sprva bela. Med razvojem v procesu nastajanja različnih organov se barva zunanjega ovoja lutke spremeni v naslednjem zaporedju: dan po mladiču so sestavljene oči še čisto bele, po 2 dneh se pojavi rumenkast odtenek, na 14. dan postanejo rožnate barve, do 16. dne pa temno vijolične.
Sklepi nog so najprej rumenkasti, od 18. dneva sklepi in kremplji na nogah pridobijo rumeno-rjavo barvo, pete in ustni deli so nekoliko svetlejši. Prsi postanejo slonokoščene. Od 19. dne začne prsni koš rahlo temneti. Trebuh slonovine. Konci segmentov nog so rjavi. Izrazitejša potemnitev je opazna na koncu brade in na dnu zgornje čeljusti. Oči postanejo vijolične. 20. dan je barva celotne lutke temno siva.
Imago oder. Na koncu stadija lutke odpade koža lutke, izpod katere se pojavi popolnoma oblikovana čebela. Čebela z mandibulami pregrizne pokrov celice in izstopi na površino satja. Novo izležena čebela ima v primerjavi s starejšo čebelo mehkejši hitinasti pokrov, poleg tega pa je njeno telo gosto poraščeno z dlačicami. Sčasoma čebele izgubijo del lasne linije, hitin pa postane veliko trši. V imaginalnem obdobju ne pride do bistvenih sprememb zunanje strukture čebeljih osebkov. "
Presnova v postembrionalnem obdobju. Ličinka in lutka sta podvrženi intenzivnim presnovnim procesom, zaradi česar se kopičijo hranila, potrebna za nastanek odrasle čebele.
Po Straussu (1911) je značilnost presnove čebel kopičenje znatnih količin glikogena kot energije za metamorfozo. Do konca faze ličinke vsebnost glikogena doseže 30% suhe mase ličinke. Hkrati se poveča količina maščobe, rast maščobnih celic.
Najprej se glukoza porabi v telesu čebel. Njegova skupna vsebnost v telesu ličink delavk je dvakrat večja kot v telesu ličink mater. Ko vsebnost glukoze v hemolimfi pade pod dovoljeno vrednost, se kompleksni ogljikovi hidrati začnejo razgrajevati in uporabljati. Najvišjo koncentracijo glukoze opazimo sredi odprte faze.
Opozoriti je treba, da je aktivnost večine proučevanih encimov največja pri mladih ličinkah. Nato se zmanjša in doseže minimum v fazi pupa. Hkrati je razvoj encimske aktivnosti pri čebelah delavkah pomembnejši kot pri maticah. Obstaja mnenje, da imajo matice v obdobju postembrionalnega razvoja bolj aerobno presnovo ogljikovih hidratov v primerjavi z delovnimi posamezniki, pospešen razvoj kraljic pa je razložen z večjo aktivnostjo številnih encimov. Razlike v količini in kakovosti mitohondrijev (celičnih organelov) pri materničnih ličinkah, starejših od
- en dan v primerjavi z ličinkami čebele delavke enake starosti. V skladu s tem opazimo tudi nekatere razlike v nekaterih oksidativnih procesih, ki potekajo v mitohondrijih.
Pri ličinki matice gre krivulja porabe kisika navzgor, dokler celica ni zaprta. Vendar pa po zapiranju krivulja privzema kisika pade (kar sovpada z vrtenjem kokona) in doseže minimum v začetni fazi lutke. Raven porabe kisika na enoto mase je pri maticah najnižja v primerjavi s čebelami delavkami in troti.
Količina absorbiranega kisika na osebka je pri dronih v postembrionalnem obdobju večja kot pri maticah in čebelah delavkah.
Dihalni koeficient, ki označuje substrat, ki se uporablja kot vir energije v celotni fazi razvoja ličinke čebele delavke, se spreminja in njegova vrednost presega enoto. Če je vrednost respiratornega koeficienta višja od ena, potem to kaže na intenzivnost procesov, povezanih s prehodom spojin, bogatih s kisikom, v spojine, revnejše s kisikom.
Povečanje respiratornega koeficienta v drugem obdobju razvoja ličink sovpada s časom prehoda na kakovostno drugačno vrsto hrane, predvsem ogljikove hidrate. Po zapiranju celice in do zabubitve ličink je respiratorni koeficient nastavljen na 0,9.
Glavni vir energije v zaprti ličinki in predkubici so ogljikovi hidrati, ki se nalagajo v obliki glikogena, v drugi polovici stadija lutke pa maščobe.
V maternici se za razliko od čebele delavke respiratorni koeficient zmanjša do 2. dneva razvoja ličink, kar je značilno za presnovo maščob. 4-5 dan iste faze je respiratorni koeficient skoraj enak vrednosti respiratornega koeficienta ličink čebele delavke, kar kaže na pretvorbo ogljikovih hidratov v maščobo. V zaprti fazi ličinke je respiratorni kvocient pod ena, kar kaže na razgradnjo ogljikovih hidratov.
Iz zgoraj navedenega lahko sklepamo, da je za izmenjavo plinov pri čebelah značilna vrsta vzorcev, značilnih za žuželke. Med razvojem ličink se intenzivnost dihanja, izražena kot količina porabljenega kisika na enoto mase, zmanjša. Vendar pa se količina kisika, porabljenega na ličinko, poveča in doseže največ pred mladičem. Na stopnji lutke se izmenjava plinov zmanjša, povečuje pa se proti izhodu odrasle osebe. Hkrati pa poleg podobnosti obstajajo tudi razlike v dihalni izmenjavi treh oblik medonosne čebele.
Pri žuželkah je razvoj razdeljen na dve obdobji - embrionalno in postembrionalno.
Embrionalni razvoj. Jajčne celice žuželk so večinoma ovalne oblike in zelo bogate z rumenjakom, ki zapolni ves osrednji del jajčeca. V skladu s tem je drobljenje jajčeca delno, površinsko; posledično nastane neprekinjena plast enakih celic, blastoderma, ki pokriva osrednji rumenjak. Hkrati del jeder ostane v masi rumenjaka: v prihodnosti se na njihov račun oblikujejo posebne celice rumenjaka. Na bodoči ventralni strani zarodka se blastoderm odebeli. To območje imenujemo zarodna črta; v prihodnosti se na njegov račun oblikuje glavni del telesa zarodka.
V predelu zarodnega traku se začnejo procesi diferenciacije. Zaradi razmnoževanja in prodiranja v globino celic blastoderme nastaneta dva trakova celic - mezodermalni trakovi, sam blastoderm pa postane ektoderm. Kasneje se mezodermalne proge razdelijo v parne primordije, kar povzroči dvojno vrsto kolomičnih vrečk. Pri višjih žuželkah (neg. Diptera) je segmentacija mezodermalnih trakov potlačena in ne pride do ločitve neodvisnih kolomov.
Vprašanje nastanka endoderme pri žuželkah še ni v celoti rešeno. V mnogih oblikah se zdi, da so rumenične celice zametek endoderma. pri Diptera in pri nekaterih drugih žuželkah sta na sprednjem in zadnjem koncu zarodnega traku ločeni dve skupini celic, ki se štejeta tudi za začetke endoderme.
Že na samem začetku polaganja zarodnih listov se začne proces nastajanja embrionalnih membran, ki je izjemno značilen za razvoj žuželk.
V najpreprostejšem in najbolj razširjenem primeru vzdolž oboda zarodnega traku ektoderm najprej oblikuje valj, nato pa dvoslojno gubo, ki postopoma narašča z vseh strani do zarodnega traku, ki je hkrati nekoliko potopljen v rumenjak. Nazadnje se robovi gube zaprejo nad zametkom zarodka, med seboj in njim pa ostane amnijska votlina, ki jo zaseda tekočina, ločena od zunanjega okolja z dvojno membrano. Zunanja plast membrane nastane kot posledica zlitja zunanjih plasti ektodermalne gube, ki se zapira nad zarodkom, in se imenuje seroza, notranja plast iz notranje plasti iste gube, slednja neposredno omejuje amnijsko votlino. in se imenuje amnion. Seroza in amnion ščitita zarodek pred poškodbami.
Medtem se zarodni trak segmentira in segmenti so izolirani, kot pri drugih členonožci in prstani, v vrstnem redu od spredaj nazaj. V odseku glave so položeni oftalmični in antenski režnji z ustreznimi rudimenti, interkalarni segment, ki se kasneje zmanjša, in 3 segmenti ustnih okončin. Nadalje posteriorno se razlikujejo 3 torakalni, ki nosijo rudimente okončin, in 11 trebušnih segmentov.
Kar zadeva diferenciacijo notranjih organov, je v svojih glavnih značilnostih naslednje. Sprednje in zadnje črevo tvorita dve ektodermalni invaginaciji. Bolj zapleteno je vprašanje izvora srednjega črevesa. Za žuželke je značilna neverjetna raznolikost načinov in virov nastanka slednjih. V nekaterih primerih se tvori zaradi endodermalnih celic rumenjaka, v drugih pa pri nastanku srednjega črevesa sodelujejo ektodermalni elementi - posebne celice, ki so lokalizirane na slepo zaprtih koncih začetkov sprednjega in zadnjega črevesja. In končno, pri številnih vrstah (partenogenetske samice listnih uši, nekateri ihnevmoni) srednje črevo tvori le ektoderm zaradi močne rasti anlage prednjega črevesa, ki doseže zadnji konec telesa. Entodermalni elementi hkrati degenerirajo. Malpigijeve žile izhajajo iz ektoderma zadnjega črevesa. Živčni sistem je položen v obliki ventralnega grebena ektoderma, ki se nato pogrezne pod ovojnico. Koelomične vrečke razpadejo in tvorijo mešano telesno votlino - mixocoel. Ob tem sproščeni celični elementi tvorijo mišice, srce, maščobno telo in nekatere druge organe. Primarne zarodne celice se odložijo zelo zgodaj, včasih že v obdobju, ko je rumenjak obdan z eno samo plastjo blastoderma.
Zarodek raste, hranljivi rumenjak se nenehno porablja, zarodne ovojnice pa počijo, se na hrbtni strani ovijejo v obliki gub, ki se degenerirajo. Zarodek je pripravljen na izhod iz jajčeca in s tem se embrionalni razvoj žuželke konča.
Postembrionalni razvoj. Pri žuželkah se iz jajčeca pojavi mlada žival ali ličinka s polnim številom segmentov. Edina izjema so primitivni kriptomaksilariji, ki pripadajo neg. Protura, pri katerem se mladiči od odraslih razlikujejo po odsotnosti treh posteriornih trebušnih segmentov. Slednji se oblikujejo iz zadnje rastne cone zaporedno po prvih moltih. Tako je za žuželke značilen razvoj brez anamorfoze, le pri proturu pa najdemo rezidualno anamorfozo.
Za postembrionalni razvoj žuželk je značilna velika raznolikost, ki jo v veliki meri določajo biološke značilnosti predstavnikov te skupine živali.
V najbolj primitivnih oblikah (podkl. Entognatha) rast in razvoj mladega organizma ne spremljajo bistvene strukturne spremembe. Mladiči so v vseh glavnih značilnostih podobni materinemu organizmu, to je, da sta ličinka in metamorfoza odsotna. V bistvu gre v tem primeru za neposredni razvoj. Krilate žuželke, ki so v evolucijskem smislu višje, imajo, nasprotno, metamorfozo, glede na naravo transformacije pa jih delimo na hemimetabolične - z nepopolno transformacijo in holometabolične - s popolno metamorfozo. Nepopolna preobrazba je značilna za nižje organizirane skupine krilatih žuželk: pravokrilce, ščurke, žuželke, enodnevnice, kačje pastirje. Z nepopolno preobrazbo je ličinka, ki se izleže iz jajčeca, v splošnem videzu že v mnogih pogledih podobna odraslemu organizmu, od njega se razlikuje predvsem v embrionalnem stanju kril, nerazvitosti sekundarnih spolnih značilnosti in pogosto prisotnosti nekaterih začasnih , čisto larvalni organi: na primer sapnične škrge pri ličinkah majske muhe itd. Krila so sprva le v obliki komaj vidnih zunanjih zametkov. Glajenje razlike med ličinko in odraslim organizmom se pojavi postopoma, ki ga spremljajo številne molte.
Z vsakim naslednjim taljenjem postane ličinka vse bolj podobna odrasli fazi žuželk ali odraslim osebam (imago), zametki kril se povečajo itd.
Pri nekaterih primitivnih žuželkah je število moltov zelo veliko in lahko doseže 25-30 (majice). Predstavniki drugih odredov imajo manj vrstic (4-5).

Pri holometabolnih žuželkah je razvoj drugačen. Ličinka se močno razlikuje od odrasle živali: sploh nima zunanjih zametkov kril, pogosto ima različno število okončin itd. Lastnosti ličink se ohranijo v nizu moltov, to je v celotnem obdobju ličinke. življenje. Ličinke holometaboličnih žuželk so izjemno raznolike. Ličinka se od imaga razlikuje po številnih primitivnih značilnostih: večja homonomija disekcije, odsotnost kril, šibek razvoj čutil (ni sestavljenih oči, ampak samo ocelli, antene so zelo majhne, itd.), v mnogih primerih pa tudi naprava in število udov. Zelo pogosto so ustni udi v starosti ličink razporejeni po eni vrsti (grizenje pri gosenicah metuljev), pri odraslih - drugače (sesalni ustni deli pri odraslih metuljih).
Kot je bilo že omenjeno, se struktura, značilna za ličinko, ohrani skozi celotno obdobje ličinke: transformacija se ne zgodi postopoma, ampak se spremembe, potrebne za metamorfozo, odložijo na čas zadnje taljenja ličinke. V tem obdobju pride do tako temeljnih preobrazb organizma ličinke, da postane nadaljevanje običajnih fizioloških funkcij, predvsem prehranjevanja in gibanja, oteženo, celo nemogoče. To se odraža v dejstvu, da ličinka po zadnjem moltu preide v stanje počitka - postane krizalis. Pupa predstavlja najbolj značilno stopnjo holometabolne transformacije. Bolj ko je popolno stanje mirovanja, bolje je izražena popolna preobrazba.
Individualni razvoj žuželke ali ontogenezo lahko razdelimo na 2 obdobji - razvoj znotraj jajčeca ali embrionalni in razvoj po izstopu iz jajčeca ali postembrionalni.
Večina žuželk odlaga jajca. Jajčece žuželke je celica, ki vsebuje jedro, protoplazmo, devtoplazmo ali rumenjak, potrebno za prehrano in razvoj zarodka. Poleg tega jajca žuželk pogosto vsebujejo simbiotske mikroorganizme, pridobljene od matere preko jajčnikov (t.j. transovarialno). Jajce je zunaj prekrito s horionom – lupino. Horion ima včasih izrazito mikroskulpturo, ki služi kot zanesljiv znak za razlikovanje rodov in celo vrst žuželk glede na fazo jajčeca. Pod horionom leži prava ali rumenjakova membrana jajčeca. Z nastankom horiona se mikropilu pusti odprtina, ki služi za prehod semenčic med oploditvijo. Jajce ima sprednji in zadnji konec, ki ustrezata položaju zarodka. V jajčniku je sprednji pol jajčeca usmerjen proti materini glavi, oziroma se nahajata hrbtna in trebušna stran jajčeca.
Velikost in videz jajčec sta različna. Velikost je 0,02 - 0,03 mm za listne uši in tripse, 8-10 mm za kobilice. Jajca so gladka pri listnih ušesih, rebrasta v mericah, bela, včasih opremljena s pokrovčkom, pokrita z gubami.
Oblika jajca. Ovalni - hrošči, metulji - podolgovati, polkrogli - zajemalke, v obliki steklenice - dnevni metulji, v obliki soda - hrošči, imajo steblo - čipke, sesalci. Odlaganje jajc: posamično, v skupinah, odprto ali v substratu, lahko zaščiteno, prekrito z izločki gonad.
Embrionalni razvoj se začne z razdrobljenostjo jedra in premikanjem nastalih hčerinskih jeder z majhnimi območji protoplazme na obrobje jajčeca. Tu iz mase hčerinskih jeder nastane neprekinjena plast celic - blastoderma. Kasneje se blastoderm diferencira na germinalno in ekstraembrionalno cono: celice slednje ne sodelujejo pri nastanku zarodka, medtem ko se celice germinalne cone začnejo intenzivneje deliti in tvorijo embrionalni pas na ventralni strani. jajčeca, ki nato štrli in nastaneta ektoderm in mezoderm.
Razvoj zarodka spremlja blastokineza: nastanejo embrionalne membrane, ločijo se segmenti glave, torakalni in trebušni del. Temelji notranje strukture se začnejo postavljati s tvorbo mezoderma. Iz ektoderma izhajajo vsi zunanji pokrovi - nato usta in anus, sprednje in zadnje črevo. Mezoderm povzroča mišični sistem, maščobno telo, hrbtno žilo, ovoj spolnih žlez.
Zarodek se začne premikati, zajame zrak, tekočino iz jajčeca, horion se zlomi in zarodek, ki se je spremenil v ličinko, pride ven.
2. Razmnoževanje žuželk ima številne značilnosti, ki so bistveni vidik njihove biologije in jih je treba upoštevati. Te značilnosti vključujejo metode razmnoževanja, dodatno prehrano, srečanje spolov in oploditev, spolno produktivnost.
Metode razmnoževanja .
Razmnoževanje pri večini žuželk spremljata parjenje in oploditev, tj. povezana z udeležbo dveh spolov - zato se imenuje biseksualna (biseksualna) gamogenetika. Večina žuželk odlaga jajca, obstajajo pa tudi drugi načini razmnoževanja: živorojenost, partenogeneza, pedogeneza, poliembrionija. Živo rojstvo je, da se embrionalni razvoj zaključi v telesu matere, zato se namesto odlaganja jajčec na svet rodijo ličinke (listne uši, gadfly, krvosese muhe). Ličinke se izležejo v jajčnih ceveh ali med prehodom skozi jajcevod.
Partenogeneza - za deviško razmnoževanje je značilna odsotnost oploditve in jo opazimo tako pri oviparnih kot pri živorodnih žuželkah.
Oblike partenogeneze:
1. arhenotokija- iz neoplojenih jajčec se razvijejo samo samci; 2) thelytokia - iz neoplojenih jajčec se razvijejo le samice; 3) amfitokija- samci in samice se razvijejo iz neoplojenih jajčec.
Partenogeneza je lahko fakultativna, stalna in ciklična.
Fakultativna partenogeneza ali spadic se manifestira nedosledno, nastane pod zunanjimi vplivi ali ob določenem fiziološkem stanju samice, ki odlaga jajčeca (maški molj, topolov jastreb).
Za družbene žuželke je značilna stalna partenogeneza arrenotokijskega tipa. Pri čebelah se iz neoplojenih jajčec vedno razvijejo samci, iz oplojenih pa samice. Pri žuželkah, ki nimajo kolonialnega načina življenja, opazimo trajno partenogenezo telitokijskega tipa. Torej pri mnogih črvih, tripsih, žagah in nekaterih jahačih samcev ni, razmnoževanje vedno poteka s partenogenezo in iz neoplojenih jajčec se izležejo samo samice.
Partenogeneza igra zelo pomembno vlogo v življenju žuželk. Zahvaljujoč partenogenezi in njenim modifikacijam se reproduktivni potencial vrste dramatično poveča: s partenogenetskim razmnoževanjem se možnosti preživetja vrste v neugodnih razmerah in med naseljevanjem znatno povečajo.
3. Po zapustitvi jajčeca se začne postembrionalni razvoj žuželke. To obdobje ni preprosta rast in povečanje telesne velikosti, temveč je značilen prehod telesa iz ene faze v drugo. Ta vrsta individualnega razvoja se imenuje metamorfoza ali razvoj s preobrazbo.
Bistvo metamorfoze je v tem, da se posameznik v razvoju skozi življenje bistveno prestrukturira v svoji morfološki organizaciji in značilnostih biologije. V zvezi s tem obstaja diferenciacija postembrionalnega razvoja v 2 fazi - ličinke in odrasle. V larvalni fazi poteka rast in razvoj osebkov, v imago fazi razmnoževanje in naselitev. V drugih primerih se med temi fazami pojavi vmesna faza - pupa. Obstajata dve glavni vrsti metamorfoze - nepopolna in popolna preobrazba. nepopolna pretvorba oz hemimetamorfoza 3 faze - jajce, ličinka, odrasel. Ličinke so podobne odraslim (sestavljene oči, ustni deli, zametki kril, podoben način življenja), podobne imagu. S popolno (holometamorfoza) 4 faze. Ličinke sploh niso videti kot odrasli, ni zapletenih oči, kril, živijo v različnih pogojih.
Nekateri imajo trebušne noge, drugačen ustni aparat, drugi imajo svilene ali pajkove žleze.
Spremembe nepopolne preobrazbe: a) hipomorfoza - poenostavljena nepopolna preobrazba in je značilna za krilate žuželke, ki so izgubile krila (sekundarne brezkrilne uši, uši, brezkrilni predstavniki kobilic, kobilice, črički, ščurki, paličnjaki, senojedci, stenice); b) hipermorfoza - zapleten nepopoln cikel (alevrodidi, resarji, kokcidni samci). Pojav na koncu faze ličink v stanju mirovanja.
Anamorfoza- ličinke so podobne odraslim, vendar imajo manjše število trebušnih segmentov; ko se ličinka razvija, zrastejo dodatni segmenti na vrhu trebuha, vendar dosežejo polno število šele v odrasli fazi.
protomorfoza– taljenje v odrasli dobi, nekaj podobnosti ličinke z odraslo fazo, vendar odsotnost delitve ličinke na prsni koš in trebuh (podura, dvorepa, ščetinasta).
hipermetamorfoza- zaplet popolne preobrazbe (prisotnost 2 oblik ličink in včasih lutk (mehurji)).
4. Faza ličink je takoj po izvalitvi iz jajčeca. Sprva svetlo, po izvalitvi je ličinka brezbarvna, ima mehke pokrove. Pri odprto živečih žuželkah ličinka hitro potemni in strdi. Prehranjevalni dražljaji nastanejo po prebavi ostankov embrionalnega rumenjaka in izločanju iztrebkov. Ličinka vstopi v par izboljšane prehrane, rasti in razvoja. Njihovo rast in razvoj spremljajo občasne taline, odpadanje kožne kožice – zaradi taljenja se telo poveča in pride do velikih sprememb. Število povezav se spreminja 3 (muhe), 4-5 (hrošči), 25-30 (padenki).
Po vsakem moltu ličinka preide v naslednjo stopnjo ali starost, zato si molti delijo starost ličink. V skladu s številom moltov se ugotavlja tudi število ličink. Določanje starosti moltov (potrebno za načrtovanje časa boja) se izvaja glede na naslednje značilnosti: antene pri kobilicah, povečanje velikosti telesa (velikost širine kapsule glave) pri gosenicah metuljev in ličinkah hroščev. . Ličinke so dveh vrst - odrasle (primarne) in sekundarne.
Naiads - ličinke s škrgami, živijo v vodi (kačji pastirji, padyonki, kamenjarji).
Campodeoid - mobilen, temno obarvan z gosto ovojnico in s 3 pari prsnih nog, dobro izolirano prognatično glavo (hroščki, plavalci, čipkarice).
Črvast - sedeč, svetle barve, brez trebušnih nog.
Gosenice imajo dobro ločeno kožo glave, 3 pare prsnih nog, 2-8 parov trebušnih.
pupalna faza. Ta faza razvoja metamorfoze je značilna samo za žuželke s popolnim ciklom. Značilnost lutke je nezmožnost hranjenja in zelo pogosto ostane nepremična. Živi od zalog, ki jih nabere ličinka, in se pogosto obravnava kot faza mirovanja.
Navzven lutka ni videti kot odrasla oseba, vendar ima številne znake odrasle faze: zunanje zametke kril, nog, antene, sestavljene oči itd. V fazi lutke pride do histolize in histogeneze.
Histoliza- razpad notranjih organov ličinke, ki ga spremlja prodiranje in prodiranje v tkiva krvnih celic - hemocitov. Hemociti delujejo kot jedci celic, tj. fagocitov, katerih povečana aktivnost vodi do uničenja in absorpcije tkivne snovi. Histoliza zajame mišični sistem, zato prepupa postane nepremična, vpliva na prebavni sistem, vendar ne vpliva na živčni in reproduktivni sistem, pa tudi na hrbtno žilo.
Histogeneza- proces nastajanja tkiv in organov zamišljenega življenja. Vir za nastanek teh novih tkiv in organov so produkti histolize. Histogeneza zajame mišični in prebavni sistem ter ju preuredi za nove, namišljene funkcije. Mišični sistem je prenovljen za nove oblike gibanja za let, prebavni sistem za nove vrste hrane.
Med histogenezo imajo glavno vlogo imaginarni rudimenti - skupine hipodermalnih celic, iz katerih izhajajo določena tkiva in organi.
Hormon taljenja je eksidon, ki je potreben za normalen razvoj ličink.
Obstajajo naslednje vrste lutk:
odprte lutke- prosti, samo na telo stisnjeni namišljeni prirastki (antene, noge, krila), značilni za hrošče.
Mladički s premičnimi mandibulami - za izhod iz kokona uporabljajo premične zgornje čeljusti in se lahko sami premikajo (mrežasti tulci, tulci, zobati molji). Mladički z negibnimi mandibulami le-teh ne morejo uporabljati, ko izstopijo iz kokona (hrošči, Hymenoptera, Fanoptera, številni Diptera).
Skrite lutke prekrita z otrdelo kožo ličink, ki ne odpada, ki igra vlogo lupine ali lažnega kokona - puparije (muhe).
pokrito- imajo namišljene dodatke, ki so tesno stisnjeni in spajkani na telo zaradi dejstva, da med zadnjim moltom ličinka izloča skrivnost, ki, ko se strdi, pokrije lutko s trdo lupino (metulji, pikapolonice).
Pred zabubitvijo se nekatere ličinke obdajo s kokonom (svila, pajčevina). Včasih je mesto mladičev rastlinska stebla, zemeljska zibelka, odprta mladica.
Imago faza.
Žuželke, ki izhajajo iz lutke, imajo znake odrasle osebe, vendar sprva krila ostanejo zložena. Nato razširijo krila, ovojnica se zgosti in pridobi barvo popolnoma oblikovane odrasle žuželke. V odrasli fazi se žuželke ne talijo in niso sposobne rasti. Izjeme so padyonki, bedaki. Biološka funkcija odrasle faze je razpršitev in razmnoževanje. To je že funkcija življenja vrste in je namenjena ohranjanju obstoja vrste. Zahvaljujoč krilom se sposobnost odraslih žuželk, da se naselijo, močno poveča, razmnoževanje pa vam omogoča, da pustite potomce na novem mestu.
Razpršitev odraslih žuželk poteka z aktivnim in pasivnim letom. Aktivni leti so običajno značilni za velike žuželke in jih opazimo pri številnih vrstah kačjih pastirjev, kobilic, metuljev, hroščev in so zelo razširjeni. Za listne uši in muhe so značilni pasivni leti.
Prehod v odrasle osebe spremljajo zunanje spremembe v barvi telesa, povečanje velikosti trebuha pri samici zaradi razvoja jajčecev, napolnjenih z jajci - pri oplojenih samicah termitov in mravelj - spuščanje kril, nezrela kobilica - roza , zrelo - svetlo rumeno.
Spolni dimorfizem se razlikuje med samci in samicami v številnih zunanjih, sekundarnih spolnih značilnostih - v obliki in velikosti anten, velikosti telesa, različnih strukturnih podrobnostih (hrošč nosorog, hrošč jelen). Moški so bolj mobilni, vodijo bolj odprt življenjski slog.
Polimorfizem - obstoj žuželk, navzven različnih oblik iste vrste - (mravlje, čebele, termiti, samci, samice, delavci, vojaki). Spolni polimorfizem je nadzorovan znotraj družine in ni odvisen od vpliva zunanjih dejavnikov.
Ekološki polimorfizem nastane pod vplivom zunanjega okolja (dolgokrili, kratkokrili, brezkrilni hrošči).